
In biology, a spore is a unit of sexual or asexual reproduction that may be adapted for dispersal and for survival, often for extended periods of time, in unfavourable conditions. Spores form part of the life cycles of many plants, algae, fungi and protozoa. They were thought to have appeared as early as the mid-late Ordovician period as an adaptation of early land plants.

A sporangium ; pl.: sporangia) is an enclosure in which spores are formed. It can be composed of a single cell or can be multicellular. Virtually all plants, fungi, and many other groups form sporangia at some point in their life cycle. Sporangia can produce spores by mitosis, but in land plants and many fungi, sporangia produce genetically distinct haploid spores by meiosis.
Chytridiomycota are a division of zoosporic organisms in the kingdom Fungi, informally known as chytrids. The name is derived from the Ancient Greek χυτρίδιον (khutrídion), meaning "little pot", describing the structure containing unreleased zoospores. Chytrids are one of the earliest diverging fungal lineages, and their membership in kingdom Fungi is demonstrated with chitin cell walls, a posterior whiplash flagellum, absorptive nutrition, use of glycogen as an energy storage compound, and synthesis of lysine by the α-amino adipic acid (AAA) pathway.

Synchytrium endobioticum is a chytrid fungus that causes the potato wart disease, or black scab. It also infects some other plants of the genus Solanum, though potato is the only cultivated host.

Peronosporaceae are a family of water moulds that contains 21 genera, comprising more than 600 species. Most of them are called downy mildews.

Phytophthora palmivora is an oomycete that causes bud-rot of palms, fruit-rot or kole-roga of coconut and areca nut. These are among the most serious diseases caused by fungi and moulds in South India. It occurs almost every year in Malnad, Mysore, North & South Kanara, Malabar and other areas. Similar diseases of palms are also known to occur in Sri Lanka, Mauritius, and Sumatra. The causative organism was first identified as P. palmivora by Edwin John Butler in 1917.
This is a glossary of some of the terms used in phytopathology.
Sporogenesis is the production of spores in biology. The term is also used to refer to the process of reproduction via spores. Reproductive spores were found to be formed in eukaryotic organisms, such as plants, algae and fungi, during their normal reproductive life cycle. Dormant spores are formed, for example by certain fungi and algae, primarily in response to unfavorable growing conditions. Most eukaryotic spores are haploid and form through cell division, though some types are diploid sor dikaryons and form through cell fusion.we can also say this type of reproduction as single pollination
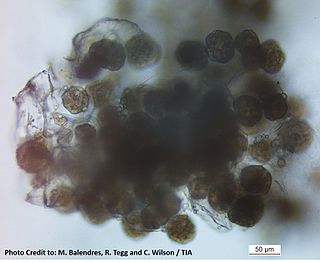
Powdery scab is a disease of potato tubers. It is caused by the cercozoan Spongospora subterranea f. sp. subterranea and is widespread in potato growing countries. Symptoms of powdery scab include small lesions in the early stages of the disease, progressing to raised pustules containing a powdery mass. These can eventually rupture within the tuber periderm. The powdery pustules contain resting spores that release anisokont zoospores to infect the root hairs of potatoes or tomatoes. Powdery scab is a cosmetic defect on tubers, which can result in the rejection of these potatoes. Potatoes which have been infected can be peeled to remove the infected skin and the remaining inside of the potato can be cooked and eaten.
Pythium irregulare is a soil borne oomycete plant pathogen. Oomycetes, also known as "water molds", are fungal-like protists. They are fungal-like because of their similar life cycles, but differ in that the resting stage is diploid, they have coenocytic hyphae, a larger genome, cellulose in their cell walls instead of chitin, and contain zoospores and oospores.

Albugo is a genus of plant-parasitic oomycetes. Those are not true fungi (Eumycota), although many discussions of this organism still treat it as a fungus. The taxonomy of this genus is incomplete, but several species are plant pathogens. Albugo is one of three genera currently described in the family Albuginaceae, the taxonomy of many species is still in flux.

Rhizophydiales are an important group of chytrid fungi. They are found in soil as well as marine and fresh water habitats where they function as parasites and decomposers.

Blastocladiomycota is one of the currently recognized phyla within the kingdom Fungi. Blastocladiomycota was originally the order Blastocladiales within the phylum Chytridiomycota until molecular and zoospore ultrastructural characters were used to demonstrate it was not monophyletic with Chytridiomycota. The order was first erected by Petersen for a single genus, Blastocladia, which was originally considered a member of the oomycetes. Accordingly, members of Blastocladiomycota are often referred to colloquially as "chytrids." However, some feel "chytrid" should refer only to members of Chytridiomycota. Thus, members of Blastocladiomycota are commonly called "blastoclads" by mycologists. Alternatively, members of Blastocladiomycota, Chytridiomycota, and Neocallimastigomycota lumped together as the zoosporic true fungi. Blastocladiomycota contains 5 families and approximately 12 genera. This early diverging branch of kingdom Fungi is the first to exhibit alternation of generations. As well, two (once) popular model organisms—Allomyces macrogynus and Blastocladiella emersonii—belong to this phylum.

Ceratiomyxa is a genus of plasmodial slime mould within the Eumycetozoa, first described by Pier Antonio Micheli. They are widely distributed and commonly found on decaying wood.

Entorrhizomycetes is the sole class in the phylum Entorrhizomycota, within the Fungi subkingdom Dikarya along with Basidiomycota and Ascomycota. It contains three genera and is a small group of teliosporic root parasites that form galls on plants in the Juncaceae (rush) and Cyperaceae (sedge) families. Prior to 2015 this phylum was placed under the subdivision Ustilaginomycotina. A 2015 study did a "comprehensive five-gene analyses" of Entorrhiza and concluded that the former class Entorrhizomycetes is possibly either a close sister group to the rest of Dikarya or Basidiomycota.
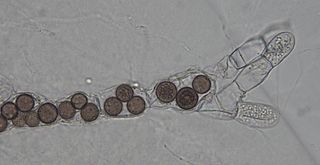
Rozella is a fungal genus of obligate endoparasites of a variety of hosts, including Oomycota, Chytridiomycota, and Blastocladiomycota. Rozella was circumscribed by French mycologist Marie Maxime Cornu in 1872. Considered one of the earliest diverging lineages of fungi, the widespread genus contains 27 species, with the most well studied being Rozella allomycis. Rozella is a member of a large clade of fungi referred to as the Cryptomycota/Rozellomycota. While some can be maintained in dual culture with the host, most have not been cultured, but they have been detected, using molecular techniques, in soil samples, and in freshwater and marine ecosystems. Zoospores have been observed, along with cysts, and the cells of some species are attached to diatoms.

Parvilucifera is a genus of marine alveolates that behave as endoparasites of dinoflagellates. It was described in 1999 by biologists Fredrik Norén and Øjvind Moestrup, who identified the genus among collections of Dinophysis dinoflagellates off the coast of Sweden. Initially mistaken for products of sexual reproduction, the round bodies found within these collections were eventually recognized as sporangia, spherical structures that generate zoospores of a parasitic protist. This organism was later identified as P. infectans, the type species. The examination of this organism and its close genetic relationship to Perkinsus led to the creation of the Perkinsozoa phylum within the Alveolata group.
Allomyces is a genus of fungi in the family Blastocladiaceae. It was circumscribed by British mycologist Edwin John Butler in 1911. Species in the genus have a polycentric thallus and reproduce sexually or asexually by zoospores that have a whiplash-like flagella. They are mostly isolated from soils in tropical countries, commonly in ponds, rice fields, and slow-moving rivers.

Physoderma is a genus of chytrid fungi. Described by German botanist Karl Friedrich Wilhelm Wallroth in 1833, the genus contains some species that are parasitic on vascular plants, including P. alfalfae and P. maydis, causative agents of crown wart of alfalfa and brown spot of corn, respectively. Of the chytrid genera, Physoderma is the oldest. However, species were confused with the rust fungi, the genus Synchytrium, and the genus Protomyces of Ascomycota. Members of Physoderma are obligate parasites of pteridophytes and angiosperms. There are approximately 80 species within this genus.
Black rot on orchids is caused by Pythium and Phytophthora species. Black rot targets a variety of orchids but Cattleya orchids are especially susceptible. Pythium ultimum and Phytophthora cactorum are known to cause black rot in orchids.
