
A hypha is a long, branching, filamentous structure of a fungus, oomycete, or actinobacterium. In most fungi, hyphae are the main mode of vegetative growth, and are collectively called a mycelium.

A mycorrhiza is a symbiotic association between a fungus and a plant. The term mycorrhiza refers to the role of the fungus in the plant's rhizosphere, its root system. Mycorrhizae play important roles in plant nutrition, soil biology, and soil chemistry.

Zygomycota, or zygote fungi, is a former division or phylum of the kingdom Fungi. The members are now part of two phyla: the Mucoromycota and Zoopagomycota. Approximately 1060 species are known. They are mostly terrestrial in habitat, living in soil or on decaying plant or animal material. Some are parasites of plants, insects, and small animals, while others form symbiotic relationships with plants. Zygomycete hyphae may be coenocytic, forming septa only where gametes are formed or to wall off dead hyphae. Zygomycota is no longer recognised as it was not believed to be truly monophyletic.

An arbuscular mycorrhiza (AM) is a type of mycorrhiza in which the symbiont fungus penetrates the cortical cells of the roots of a vascular plant forming arbuscules. Arbuscular mycorrhiza is a type of endomycorrhiza along with ericoid mycorrhiza and orchid mycorrhiza .They are characterized by the formation of unique tree-like structures, the arbuscules. In addition, globular storage structures called vesicles are often encountered.
Glomeromycota are one of eight currently recognized divisions within the kingdom Fungi, with approximately 230 described species. Members of the Glomeromycota form arbuscular mycorrhizas (AMs) with the thalli of bryophytes and the roots of vascular land plants. Not all species have been shown to form AMs, and one, Geosiphon pyriformis, is known not to do so. Instead, it forms an endocytobiotic association with Nostoc cyanobacteria. The majority of evidence shows that the Glomeromycota are dependent on land plants for carbon and energy, but there is recent circumstantial evidence that some species may be able to lead an independent existence. The arbuscular mycorrhizal species are terrestrial and widely distributed in soils worldwide where they form symbioses with the roots of the majority of plant species (>80%). They can also be found in wetlands, including salt-marshes, and associated with epiphytic plants.

The Mucorales is the largest and best-studied order of zygomycete fungi. Members of this order are sometimes called pin molds. The term mucormycosis is now preferred for infections caused by molds belonging to the order Mucorales.

Glomus is a genus of arbuscular mycorrhizal (AM) fungi, and all species form symbiotic relationships (mycorrhizae) with plant roots. Glomus is the largest genus of AM fungi, with ca. 85 species described, but is currently defined as non-monophyletic.

Mucor is a microbial genus of approximately 40 species of molds in the family Mucoraceae. Species are commonly found in soil, digestive systems, plant surfaces, some cheeses like Tomme de Savoie, rotten vegetable matter and iron oxide residue in the biosorption process.

Mortierella species are soil fungi belonging to the order Mortierellales within the subphylum Mortierellomycotina. The widespread genus contains about 85 species.
Mucor mucedo, commonly known as the common pinmould, is a fungal plant pathogen and member of the phylum Mucoromycota and the genus Mucor. Commonly found on soil, dung, water, plants and moist foods, Mucor mucedo is a saprotrophic fungus found world-wide with 85 known strains. It is often mistaken for Rhizopus rots on fruits due to similar mould growth shape and colour. Contrastingly, however, Mucor mucedo is found to grow on a wide range of stored grains and plants, including cucumber and tomato. Discovered in Italy in 1729 by P.A. Micheli and later noted by Carl Linnaeus in 1753 in the Species Plantarum, Mucor mucedo was originally classified as Mucor vulgaris by Micheli but later classified synonymous under name Mucor mucedo. The species was redescribed as Ascophora mucedo by H.J. Tode in 1790 but this type resided in a stoloniferous habitat and was later made the type of new genus Rhizopus.

Mucor racemosus is a rapidly growing, weedy mould belonging to the division Mucoromycota. It is one of the earliest fungi to be grown in pure culture and was first isolated in 1886. It has a worldwide distribution and colonizes many habitats such as vegetational products, soil and houses. The fungus is mostly known for its ability to exhibit both filamentous and yeast-like morphologies, often referred to as dimorphism. Stark differences are seen in both forms and conditions of the environment heavily affect the phases of the M. racemosus. Like many fungi, it also reproduces both sexually and asexually. The dimorphic capacity of this species has been proposed as an important factor in its pathogenicity and has enhanced the industrial importance. This species is considered an opportunistic pathogen, generally limited to immunocompromised individuals. It also been associated with allergy and inflammations of facial sinuses. Its association with allergy has made it a common fungus used in allergen medical testing. Industrial use of the fungus is in the production of enzymes and the manufacture of certain dairy foods.

A fungus is any member of the group of eukaryotic organisms that includes microorganisms such as yeasts and molds, as well as the more familiar mushrooms. These organisms are classified as one of the traditional eukaryotic kingdoms, along with Animalia, Plantae and either Protista or Protozoa and Chromista.

Phycomycetes is an obsolete polyphyletic taxon for certain fungi with aseptate hyphae. It is used in the Engler system. Asexual reproduction takes place by zoospores (motile) or by Aplanospores (non-motile). These spores are endogenously produced in sporangium. A zygospore is formed by fusion of two gametes. These gametes are similar in morphology (isogamous) or dissimilar.

Mucoromycotina is a subphylum of uncertain placement in Fungi. It was considered part of the phylum Zygomycota, but recent phylogenetic studies have shown that it was polyphyletic and thus split into several groups, it is now thought to be a paraphyletic grouping. Mucoromycotina is currently composed of 3 orders, 61 genera, and 325 species. Some common characteristics seen throughout the species include: development of coenocytic mycelium, saprotrophic lifestyles, and filamentous.
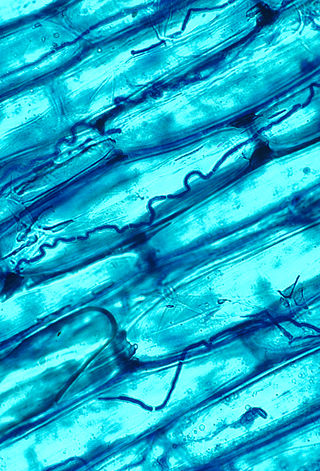
Plant use of endophytic fungi in defense occurs when endophytic fungi, which live symbiotically with the majority of plants by entering their cells, are utilized as an indirect defense against herbivores. In exchange for carbohydrate energy resources, the fungus provides benefits to the plant which can include increased water or nutrient uptake and protection from phytophagous insects, birds or mammals. Once associated, the fungi alter nutrient content of the plant and enhance or begin production of secondary metabolites. The change in chemical composition acts to deter herbivory by insects, grazing by ungulates and/or oviposition by adult insects. Endophyte-mediated defense can also be effective against pathogens and non-herbivory damage.

Rhizophagus irregularis is an arbuscular mycorrhizal fungus used as a soil inoculant in agriculture and horticulture. Rhizophagus irregularis is also commonly used in scientific studies of the effects of arbuscular mycorrhizal fungi on plant and soil improvement. Until 2001, the species was known and widely marketed as Glomus intraradices, but molecular analysis of ribosomal DNA led to the reclassification of all arbuscular fungi from Zygomycota phylum to the Glomeromycota phylum.
Dark septate endophytes (DSE) are a group of endophytic fungi characterized by their morphology of melanized, septate, hyphae. This group is likely paraphyletic, and contain conidial as well as sterile fungi that colonize roots intracellularly or intercellularly. Very little is known about the number of fungal taxa within this group, but all are in the Ascomycota. They are found in over 600 plant species and across 114 families of angiosperms and gymnosperms and co-occur with other types of mycorrhizal fungi. They have a wide global distribution and can be more abundant in stressed environments. Much of their taxonomy, physiology, and ecology are unknown.
Umbelopsis ramanniana is a common and abundant soil fungus. Although the ecological role of this fungus in natural ecosystems is not yet known, it is a cosmopolitan saprotroph in soil, forest leaf litter, in animal dung, and on the spore-producing bodies of ascomycete fungi. Umbelopsis ramanniana has also been found growing as an endophyte within xylem tissue of both healthy and declining conifers, but its exact effect on the plant hosts is unknown. Umbelopsis ramanniana is a representative of a unique group of zygomycete fungi that is distinct from the Mucoromycotina and Mortierellomycotina and instead forms an early diverging lineage within the Mucoralean fungi. Umbelopsis ramanniana is important from a biochemistry and biotechnology perspective because it is highly tolerant to fungicides of benomyl group, and it is oleaginous. Expression of Umbelopsis ramanniana diacylglycerol O-acyltransferase 2A in seed increases oil in soybean without reduction of other important yield parameters. This increase in oil can potentially add over $1 billion to the annual value of soybean crops.

Glomus macrocarpum is a vesicular-arbuscular endomycorrhizal plant pathogen in the Glomeraceae family of fungi. Also occasionally known as Endogone macrocarpa, G. macrocarpum is pathogenic to multiple plants, including tobacco and chili plants. G. macrocarpum was first discovered in the French woodlands by the Tulsane brothers in the early to mid 1800s. Their first known description of G. macrocarpum was published in the New Italian Botanical Journal in 1845. G. macrocarpum has since been documented in over 26 countries, including Australia, China, and Japan for example. G. macrocarpum is frequently found in grassy meadows, forests, greenhouses, and fruit orchards. It is known for its small, round-edged, and light brown to yellow-brown sporocarp. G. macrocarpum is sometimes known as the Glomerales truffle.

The common symbiosis signaling pathway (CSSP) is a signaling cascade in plants that allows them to interact with symbiotic microbes. It corresponds to an ancestral pathway that plants use to interact with arbuscular mycorrhizal fungi (AMF). It is known as "common" because different evolutionary younger symbioses also use this pathway, notably the root nodule symbiosis with nitrogen-fixing rhizobia bacteria. The pathway is activated by both Nod-factor perception, as well as by Myc-factor perception that are released from AMF. The pathway is distinguished from the pathogen recognition pathways, but may have some common receptors involved in both pathogen recognition as well as CSSP. A recent work by Kevin Cope and colleagues showed that ectomycorrhizae also uses CSSP components such as Myc-factor recognition.


