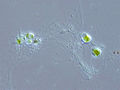
The Rhizaria are a diverse and species-rich supergroup of mostly unicellular eukaryotes. Except for the Chlorarachniophytes and three species in the genus Paulinella in the phylum Cercozoa, they are all non-photosynthethic, but many foraminifera and radiolaria have a symbiotic relationship with unicellular algae. A multicellular form, Guttulinopsis vulgaris, a cellular slime mold, has been described. This group was used by Cavalier-Smith in 2002, although the term "Rhizaria" had been long used for clades within the currently recognized taxon. Being described mainly from rDNA sequences, they vary considerably in form, having no clear morphological distinctive characters (synapomorphies), but for the most part they are amoeboids with filose, reticulose, or microtubule-supported pseudopods. In the absence of an apomorphy, the group is ill-defined, and its composition has been very fluid. Some Rhizaria possess mineral exoskeletons, which are in different clades within Rhizaria made out of opal, celestite, or calcite. Certain species can attain sizes of more than a centimeter with some species being able to form cylindrical colonies approximately 1 cm in diameter and greater than 1 m in length. They feed by capturing and engulfing prey with the extensions of their pseudopodia; forms that are symbiotic with unicellular algae contribute significantly to the total primary production of the ocean.
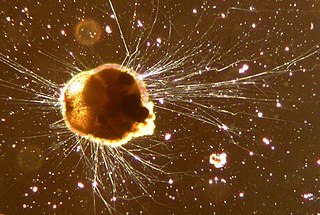
Phaeodarea or Phaeodaria is a group of amoeboid cercozoan organisms. They are traditionally considered radiolarians, but in molecular trees do not appear to be close relatives of the other groups, and are instead placed among the Cercozoa. They are distinguished by the structure of their central capsule and by the presence of a phaeodium, an aggregate of waste particles within the cell.

Cercomonads are small amoeboflagellates, widespread in aqueous habitats and common in soils.

The tectofilosids are a group of filose amoebae with shells. These are composed of organic materials and sometimes collected debris, in contrast to the euglyphids, which produce shells from siliceous scales. The shell usually has a single opening, but in Amphitrema and a few other genera it has two on opposite ends. The cell itself occupies most of the shell. They are most often found on marsh plants such as Sphagnum.
Monadofilosa is a grouping of Cercozoa. These organisms are single-celled amoeboid protists.

Telonemia is a phylum of microscopic eukaryotes commonly known as telonemids. They are unicellular free-living flagellates with a unique combination of cell structures, including a highly complex cytoskeleton unseen in other eukaryotes.
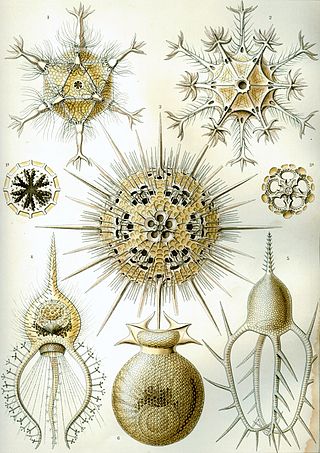
Retaria is a clade within the supergroup Rhizaria containing the Foraminifera and the Radiolaria. In 2019, the Retaria were recognized as a basal Rhizaria group, as sister of the Cercozoa.

Endomyxa is a group of eukaryotic organisms in the supergroup Rhizaria. They were initially a subphylum of Cercozoa and later a subphylum of Retaria, but several analyses have proven they are a phylogenetically separate lineage, and Endomyxa is currently regarded as its own phylum.
Thecofilosea is a class of unicellular testate amoebae belonging to the phylum Cercozoa. They are amoeboflagellates, organisms with flagella and pseudopodia, distinguished from other cercozoa by their scale-lacking test composed of organic material. They are closely related to the Imbricatea, a group of testate amoebae with tests composed of inorganic silica scales.

The spongomonads are a group of flagellated protists in the phylum Cercozoa. Taxonomically, they compose the family Sarcomonadidae and order Sarcomonadida. They were originally placed among the Reticulofilosa, but were later transferred to Monadofilosa. It includes only two genera:
The sarcomonads or class Sarcomonadea are a group of amoeboid biciliate protists in the phylum Cercozoa. They are characterized by a propensity to move through gliding on their posterior cilium or through filopodia, a lack of scales or external theca, a soft cell surface without obvious cortical filamentous or membranous skeleton, two cilia without scales or hairs, tubular mitochondrial cristae, near-spherical extrusomes, and a microbody attached to the nucleus.

Halvaria is a taxonomic grouping of protists that includes Alveolata and Stramenopiles (Heterokonta).

The vampyrellids, colloquially known as vampire amoebae, are a group of free-living predatory amoebae classified as part of the lineage Endomyxa. They are distinguished from other groups of amoebae by their irregular cell shape with propensity to fuse and split like plasmodial organisms, and their life cycle with a digestive cyst stage that digests the gathered food. They appear worldwide in marine, brackish, freshwater and soil habitats. They are important predators of an enormous variety of microscopic organisms, from algae to fungi and animals. They are also known as aconchulinid amoebae.

The plasmodiophores are a group of obligate endoparasitic protists belonging to the subphylum Endomyxa in Cercozoa. Taxonomically, they are united under a single family Plasmodiophoridae, order Plasmodiophorida, sister to the phagomyxids.
Cryptofilida is an order of small heterotrophic protists in the phylum Cercozoa. They are filose amoebae that lack cilia and gliding, and are instead characterized by movement through branching or unbranched granular filopodia that are appressed to the substrate during their feeding.

The glissomonads are a group of bacterivorous gliding flagellated protists that compose the order Glissomonadida, in the amoeboflagellate phylum Cercozoa. They comprise a vast, largely undescribed diversity of soil and freshwater organisms. They are the sister group to cercomonads; the two orders form a solid clade of gliding soil-dwelling flagellates called Pediglissa.

Euglyphia is a group of imbricate protists in the phylum Cercozoa. They are unicellular eukaryotes characterized by a cell body covered in large imbricate scales, and an apical aperture through which they extend either filose pseudopodia or two cilia of different sizes that are not used for gliding.
Pediglissa is a subclass of phagotrophic protists that inhabit soil or freshwater habitats. They were defined in 2018 according to phylogenetic analyses that showed a clade containing the orders Cercomonadida and Glissomonadida. They're the sister group of Paracercomonadida.
Helkesida is a group of microscopic protists belonging to the supergroup Rhizaria, both discovered through molecular phylogenetic analyses. It contains amoeboid flagellates with two flagella. They are either free-living, mostly on fecal matter, or live inside the gut of animals. Among these amoebae, one lineage has independently evolved aggregative multicellularity similarly to slime moulds.
Limnofila is a genus of heterotrophic protists that live in freshwater habitats and feed on bacteria. They are also present in the soil ecosystem, where they play an important role as predators of bacteria. They are classified as a single family Limnofilidae and order Limnofilida.