In biology, a kingdom is the second highest taxonomic rank, just below domain. Kingdoms are divided into smaller groups called phyla.
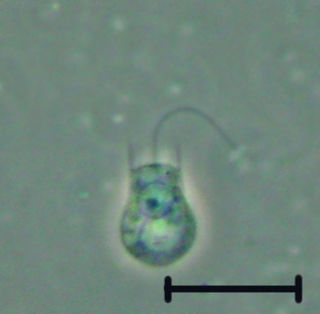
The choanoflagellates are a group of free-living unicellular and colonial flagellate eukaryotes considered to be the closest living relatives of the animals. Choanoflagellates are collared flagellates, having a funnel shaped collar of interconnected microvilli at the base of a flagellum. Choanoflagellates are capable of both asexual and sexual reproduction. They have a distinctive cell morphology characterized by an ovoid or spherical cell body 3–10 µm in diameter with a single apical flagellum surrounded by a collar of 30–40 microvilli. Movement of the flagellum creates water currents that can propel free-swimming choanoflagellates through the water column and trap bacteria and detritus against the collar of microvilli, where these foodstuffs are engulfed. This feeding provides a critical link within the global carbon cycle, linking trophic levels. In addition to their critical ecological roles, choanoflagellates are of particular interest to evolutionary biologists studying the origins of multicellularity in animals. As the closest living relatives of animals, choanoflagellates serve as a useful model for reconstructions of the last unicellular ancestor of animals.

The opisthokonts are a broad group of eukaryotes, including both the animal and fungus kingdoms. The opisthokonts, previously called the "Fungi/Metazoa group", are generally recognized as a clade. Opisthokonts together with Apusomonadida and Breviata comprise the larger clade Obazoa.

Amorphea is a taxonomic supergroup that includes the basal Amoebozoa and Obazoa. That latter contains the Opisthokonta, which includes the Fungi, Animals and the Choanomonada, or Choanoflagellates. The taxonomic affinities of the members of this clade were originally described and proposed by Thomas Cavalier-Smith in 2002.

The Ichthyosporea are a small group of Opisthokonta in Eukaryota, mostly parasites of fish and other animals.
A protist or protoctist is any eukaryotic organism that is not an animal, plant, or fungus. Protists do not form a natural group, or clade, but are a polyphyletic grouping of several independent clades that evolved from the last eukaryotic common ancestor.
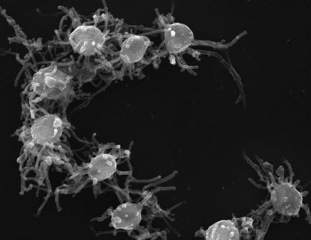
Capsaspora is a monotypic genus containing the single species Capsaspora owczarzaki. C. owczarzaki is a single-celled eukaryote that occupies a key phylogenetic position in our understanding of the origin of animal multicellularity, as one of the closest unicellular relatives to animals. It is, together with Ministeria vibrans, a member of the Filasterea clade. This amoeboid protist has been pivotal to unravel the nature of the unicellular ancestor of animals, which has been proved to be much more complex than previously thought.

Ministeria vibrans is a bacterivorous amoeba with filopodia that was originally described to be suspended by a flagellum-like stalk attached to the substrate. Molecular and experimental work later on demonstrated the stalk is indeed a flagellar apparatus.

Corallochytrium belongs to the class of Corallochytrea within Teretosporea and is a sister group to Ichthyosporea. Corallochytrium limacisporum is the only species of Corallochytrium known so far. It was first discovered and named in the Arabian Sea’s coral lagoons by Kaghu-Kumar in 1987. It was first thought to be a member of the fungi-like thraustochytrids, however, this was later disproven due to Corallochytriums lack of cilia and sagenogenetosome. Little research has been done on the life cycle or morphology. Most research concerning this genus has been done to uncover the evolution of animals and fungi, as Corallochytrium possess both animal and fungal enzymatic trademarks.
Filasterea is a proposed basal Filozoan clade of single-celled ameboid eukaryotes that includes Ministeria and Capsaspora. It is a sister clade to the Choanozoa in which the Choanoflagellatea and Animals appeared, originally proposed by Shalchian-Tabrizi et al. in 2008, based on a phylogenomic analysis with dozens of genes. Filasterea was found to be the sister-group to the clade composed of Metazoa and Choanoflagellata within the Opisthokonta, a finding that has been further corroborated with additional, more taxon-rich, phylogenetic analyses.

The apusomonads are a group of protozoan zooflagellates that glide on surfaces, and mostly consume prokaryotes. They are of particular evolutionary interest because they appear to be the sister group to the Opisthokonts, the clade that includes both animals and fungi. Together with the Breviatea, these form the Obazoa clade.

The Filozoa are a monophyletic grouping within the Opisthokonta. They include animals and their nearest unicellular relatives.

Holomycota or Nucletmycea are a basal Opisthokont clade as sister of the Holozoa. It consists of the Cristidiscoidea and the kingdom Fungi. The position of nucleariids, unicellular free-living phagotrophic amoebae, as the earliest lineage of Holomycota suggests that animals and fungi independently acquired complex multicellularity from a common unicellular ancestor and that the osmotrophic lifestyle was originated later in the divergence of this eukaryotic lineage. Opisthosporidians is a recently proposed taxonomic group that includes aphelids, Microsporidia and Cryptomycota, three groups of endoparasites.

Choanozoa is a clade of opisthokont eukaryotes consisting of the choanoflagellates (Choanoflagellatea) and the animals. The sister-group relationship between the choanoflagellates and animals has important implications for the origin of the animals. The clade was identified in 2015 by Graham Budd and Sören Jensen, who used the name Apoikozoa. The 2018 revision of the classification first proposed by the International Society of Protistologists in 2012 recommends the use of the name Choanozoa.
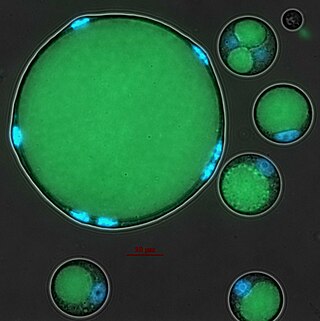
Abeoforma whisleri is a single-celled eukaryote that belongs to the Ichthyosporea clade, a group of protists closely related to animals.

Pirum gemmata is a unicellular eukaryote that belongs to the Ichthyosporea clade, a group of protists closely related to animals. P. gemmata was isolated from the gut contents of a marine invertebrate, specifically the detritivorous peanut worm Phascolosoma agassizii.
Pluriformea is a proposed sibling clade of the Filozoa, and consists of Syssomonas multiformis and the Corallochytrea. Together with the Ichthyosporea they form the Holozoa.
Tunicaraptor is a genus of marine microbial protists containing the single species Tunicaraptor unikontum, discovered in 2020 from marine waters of Chile. It is a lineage of predatorial flagellates closely related to animals. It has a rare feeding structure not seen in other opisthokonts.

Syssomonas is a monotypic genus of unicellular flagellated protists containing the species Syssomonas multiformis. It is a member of Pluriformea inside the lineage of Holozoa, a clade containing animals and their closest protistan relatives. It lives in freshwater habitats. It has a complex life cycle that includes unicellular amoeboid and flagellated phases, as well as multicellular aggregates, depending on the growth medium and nutritional state.

An amoeboflagellate is any eukaryotic organism capable of behaving as an amoeba and as a flagellate at some point during their life cycle. Amoeboflagellates present both pseudopodia and at least one flagellum, often simultaneously.