Asterales is an order of dicotyledonous flowering plants that includes the large family Asteraceae known for composite flowers made of florets, and ten families related to the Asteraceae. While asterids in general are characterized by fused petals, composite flowers consisting of many florets create the false appearance of separate petals.
Cladistics is an approach to biological classification in which organisms are categorized in groups ("clades") based on hypotheses of most recent common ancestry. The evidence for hypothesized relationships is typically shared derived characteristics (synapomorphies) that are not present in more distant groups and ancestors. However, from an empirical perspective, common ancestors are inferences based on a cladistic hypothesis of relationships of taxa whose character states can be observed. Theoretically, a last common ancestor and all its descendants constitute a (minimal) clade. Importantly, all descendants stay in their overarching ancestral clade. For example, if the terms worms or fishes were used within a strict cladistic framework, these terms would include humans. Many of these terms are normally used paraphyletically, outside of cladistics, e.g. as a 'grade', which are fruitless to precisely delineate, especially when including extinct species. Radiation results in the generation of new subclades by bifurcation, but in practice sexual hybridization may blur very closely related groupings.
In biological phylogenetics, a clade, also known as a monophyletic group or natural group, is a grouping of organisms that are monophyletic – that is, composed of a common ancestor and all its lineal descendants – on a phylogenetic tree. In the taxonomical literature, sometimes the Latin form cladus is used rather than the English form. Clades are the fundamental unit of cladistics, a modern approach to taxonomy adopted by most biological fields.

Homininae, also called "African hominids" or "African apes", is a subfamily of Hominidae. It includes two tribes, with their extant as well as extinct species: 1) the tribe Hominini ―and 2) the tribe Gorillini (gorillas). Alternatively, the genus Pan is sometimes considered to belong to its own third tribe, Panini. Homininae comprises all hominids that arose after orangutans split from the line of great apes. The Homininae cladogram has three main branches, which lead to gorillas, and to humans and chimpanzees via the tribe Hominini and subtribes Hominina and Panina. There are two living species of Panina and two living species of gorillas, but only one extant human species. Traces of extinct Homo species, including Homo floresiensis have been found with dates as recent as 40,000 years ago. Organisms in this subfamily are described as hominine or hominines.

Saxifragales is an order of angiosperms, or flowering plants, containing 15 botanical families and around 100 genera, with nearly 2,500 species. Of the 15 families, many are small, with eight of them being monotypic. The largest family is the Crassulaceae (stonecrops), a diverse group of mostly succulent plants, with about 35 genera. Saxifragales are found worldwide, primarily in temperate to subtropical zones, rarely being encountered growing wild in the tropics; however, many species are now cultivated throughout the world as knowledge of plant husbandry has improved. They can be found in a wide variety of environments, from deserts to fully aquatic habitats, with species adapted to alpine, forested or fully-aquatic habitats. Many are epiphytic or lithophytic, growing on exposed cliff faces, on trees or on rocks, and not requiring a highly organic or nutrient-dense substrate to thrive.

The Fabaceae or Leguminosae, commonly known as the legume, pea, or bean family, are a large and agriculturally important family of flowering plants. It includes trees, shrubs, and perennial or annual herbaceous plants, which are easily recognized by their fruit (legume) and their compound, stipulate leaves. The family is widely distributed, and is the third-largest land plant family in number of species, behind only the Orchidaceae and Asteraceae, with about 765 genera and nearly 20,000 known species.

A ratite is any of a group of mostly flightless birds within the infraclass Palaeognathae. They are mostly large, long-necked, and long-legged, the exception being the kiwi, which is also the only nocturnal extant ratite.

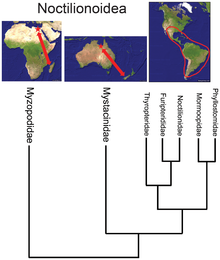
Afrotheria is a superorder of mammals, the living members of which belong to groups that are either currently living in Africa or of African origin: golden moles, elephant shrews, otter shrews, tenrecs, aardvarks, hyraxes, elephants, sea cows, and several extinct clades. Most groups of afrotheres share little or no superficial resemblance, and their similarities have only become known in recent times because of genetics and molecular studies. Many afrothere groups are found mostly or exclusively in Africa, reflecting the fact that Africa was an island continent from the Cretaceous until the early Miocene around 20 million years ago, when Afro-Arabia collided with Eurasia.

Haplorhini, the haplorhines or the "dry-nosed" primates is a suborder of primates containing the tarsiers and the simians, as sister of the Strepsirrhini ("moist-nosed"). The name is sometimes spelled Haplorrhini. The simians include catarrhines, and the platyrrhines.

In cladistics or phylogenetics, an outgroup is a more distantly related group of organisms that serves as a reference group when determining the evolutionary relationships of the ingroup, the set of organisms under study, and is distinct from sociological outgroups. The outgroup is used as a point of comparison for the ingroup and specifically allows for the phylogeny to be rooted. Because the polarity (direction) of character change can be determined only on a rooted phylogeny, the choice of outgroup is essential for understanding the evolution of traits along a phylogeny.

Pierolapithecus catalaunicus is an extinct species of primate which lived around 12.5-13 million years ago during the Miocene in what is now Hostalets de Pierola, Catalonia, Spain. Some researchers believe that it is a candidate for common ancestor to the great ape clade, or is at least closer than any previous fossil discovery. Others suggest it being a pongine, or a dryopith. On 16 October 2023, scientists reported the facial reconstruction of the great ape.

The Hominini form a taxonomic tribe of the subfamily Homininae ("hominines"). Hominini includes the extant genera Homo (humans) and Pan and in standard usage excludes the genus Gorilla (gorillas).

In the APG IV system (2016) for the classification of flowering plants, the name asterids denotes a clade. Asterids is the largest group of flowering plants, with more than 80,000 species, about a third of the total flowering plant species. Well-known plants in this clade include the common daisy, forget-me-nots, nightshades, the common sunflower, petunias, yacon, morning glory, lettuce, sweet potato, coffee, lavender, lilac, olive, jasmine, honeysuckle, ash tree, teak, snapdragon, sesame, psyllium, garden sage, table herbs such as mint, basil, and rosemary, and rainforest trees such as Brazil nut.
Phylogenetic nomenclature is a method of nomenclature for taxa in biology that uses phylogenetic definitions for taxon names as explained below. This contrasts with the traditional method, by which taxon names are defined by a type, which can be a specimen or a taxon of lower rank, and a description in words. Phylogenetic nomenclature is regulated currently by the International Code of Phylogenetic Nomenclature (PhyloCode).

The Hominidae, whose members are known as the great apes or hominids, are a taxonomic family of primates that includes eight extant species in four genera: Pongo ; Gorilla ; Pan ; and Homo, of which only modern humans remain.

Mesangiospermae is a clade of flowering plants (angiosperms), informally called "mesangiosperms". They are one of two main groups of angiosperms. It is a name created under the rules of the PhyloCode system of phylogenetic nomenclature. There are about 350,000 species of mesangiosperms. The mesangiosperms contain about 99.95% of the flowering plants, assuming that there are about 175 species not in this group and about 350,000 that are. While such a clade with a similar circumscription exists in the APG III system, it was not given a name.
The chimpanzee–human last common ancestor (CHLCA) is the last common ancestor shared by the extant Homo (human) and Pan genera of Hominini. Estimates of the divergence date vary widely from thirteen to five million years ago.

A seed plant or spermatophyte, also known as a phanerogam or a phaenogam, is any plant that produces seeds. It is a category of embryophyte that includes most of the familiar land plants, including the flowering plants and the gymnosperms, but not ferns, mosses, or algae.
The phylogenetic split of Hominidae into the subfamilies Homininae and Ponginae is dated to the middle Miocene, roughly 18 to 14 million years ago. This split is also referenced as the "orangutan–human last common ancestor" by Jeffrey H. Schwartz, professor of anthropology at the University of Pittsburgh School of Arts and Sciences, and John Grehan, director of science at the Buffalo Museum.

In phylogenetic nomenclature, the Pentapetalae are a large group of eudicots that were informally referred to as the "core eudicots" in some papers on angiosperm phylogenetics. They comprise an extremely large and diverse group accounting for about 65% of the species richness of the angiosperms, with wide variability in habit, morphology, chemistry, geographic distribution, and other attributes. Classical systematics, based solely on morphological information, was not able to recognize this group. In fact, the circumscription of the Pentapetalae as a clade is based on strong evidence obtained from DNA molecular analysis data.