Cladistics is an approach to biological classification in which organisms are categorized in groups ("clades") based on hypotheses of most recent common ancestry. The evidence for hypothesized relationships is typically shared derived characteristics (synapomorphies) that are not present in more distant groups and ancestors. However, from an empirical perspective, common ancestors are inferences based on a cladistic hypothesis of relationships of taxa whose character states can be observed. Theoretically, a last common ancestor and all its descendants constitute a (minimal) clade. Importantly, all descendants stay in their overarching ancestral clade. For example, if the terms worms or fishes were used within a strict cladistic framework, these terms would include humans. Many of these terms are normally used paraphyletically, outside of cladistics, e.g. as a 'grade', which are fruitless to precisely delineate, especially when including extinct species. Radiation results in the generation of new subclades by bifurcation, but in practice sexual hybridization may blur very closely related groupings.
In biological phylogenetics, a clade, also known as a monophyletic group or natural group, is a grouping of organisms that are monophyletic – that is, composed of a common ancestor and all its lineal descendants – on a phylogenetic tree. In the taxonomical literature, sometimes the Latin form cladus is used rather than the English form. Clades are the fundamental unit of cladistics, a modern approach to taxonomy adopted by most biological fields.

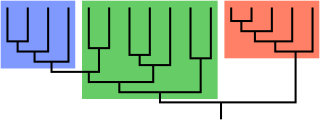
In biological cladistics for the classification of organisms, monophyly is the condition of a taxonomic grouping being a clade – that is, a grouping of taxa which meets these criteria:
- the grouping contains its own most recent common ancestor, i.e. excludes non-descendants of that common ancestor
- the grouping contains all the descendants of that common ancestor, without exception
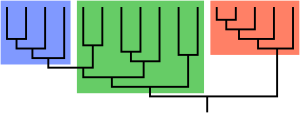
Paraphyly is a taxonomic term describing a grouping that consists of the grouping's last common ancestor and most of its descendants, but excludes one or more subgroups. The grouping is said to be paraphyletic with respect to the excluded subgroups. In contrast, a monophyletic grouping includes a common ancestor and all of its descendants.
In biology, taxonomy is the scientific study of naming, defining (circumscribing) and classifying groups of biological organisms based on shared characteristics. Organisms are grouped into taxa and these groups are given a taxonomic rank; groups of a given rank can be aggregated to form a more inclusive group of higher rank, thus creating a taxonomic hierarchy. The principal ranks in modern use are domain, kingdom, phylum, class, order, family, genus, and species. The Swedish botanist Carl Linnaeus is regarded as the founder of the current system of taxonomy, as he developed a ranked system known as Linnaean taxonomy for categorizing organisms and binomial nomenclature for naming organisms.

A cladogram is a diagram used in cladistics to show relations among organisms. A cladogram is not, however, an evolutionary tree because it does not show how ancestors are related to descendants, nor does it show how much they have changed, so many differing evolutionary trees can be consistent with the same cladogram. A cladogram uses lines that branch off in different directions ending at a clade, a group of organisms with a last common ancestor. There are many shapes of cladograms but they all have lines that branch off from other lines. The lines can be traced back to where they branch off. These branching off points represent a hypothetical ancestor which can be inferred to exhibit the traits shared among the terminal taxa above it. This hypothetical ancestor might then provide clues about the order of evolution of various features, adaptation, and other evolutionary narratives about ancestors. Although traditionally such cladograms were generated largely on the basis of morphological characters, DNA and RNA sequencing data and computational phylogenetics are now very commonly used in the generation of cladograms, either on their own or in combination with morphology.

Chromista is a proposed but polyphyletic biological kingdom consisting of single-celled and multicellular eukaryotic species that share similar features in their photosynthetic organelles (plastids). It includes all eukaryotes whose plastids contain chlorophyll c and are surrounded by four membranes. If the ancestor already possessed chloroplasts derived by endosymbiosis from red algae, all non-photosynthetic Chromista have secondarily lost the ability to photosynthesise. Its members might have arisen independently as separate evolutionary groups from the last eukaryotic common ancestor.
Evolutionary taxonomy, evolutionary systematics or Darwinian classification is a branch of biological classification that seeks to classify organisms using a combination of phylogenetic relationship, progenitor-descendant relationship, and degree of evolutionary change. This type of taxonomy may consider whole taxa rather than single species, so that groups of species can be inferred as giving rise to new groups. The concept found its most well-known form in the modern evolutionary synthesis of the early 1940s.
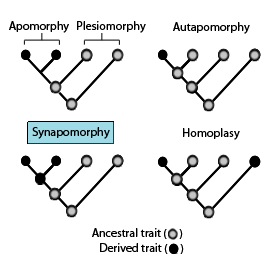
In phylogenetics, an apomorphy is a novel character or character state that has evolved from its ancestral form. A synapomorphy is an apomorphy shared by two or more taxa and is therefore hypothesized to have evolved in their most recent common ancestor. In cladistics, synapomorphy implies homology.

Lepospondyli is a diverse taxon of early tetrapods. With the exception of one late-surviving lepospondyl from the Late Permian of Morocco, lepospondyls lived from the Early Carboniferous (Mississippian) to the Early Permian and were geographically restricted to what is now Europe and North America. Five major groups of lepospondyls are known: Adelospondyli; Aïstopoda; Lysorophia; Microsauria; and Nectridea. Lepospondyls have a diverse range of body forms and include species with newt-like, eel- or snake-like, and lizard-like forms. Various species were aquatic, semiaquatic, or terrestrial. None were large, and they are assumed to have lived in specialized ecological niches not taken by the more numerous temnospondyl amphibians that coexisted with them in the Paleozoic. Lepospondyli was named in 1888 by Karl Alfred von Zittel, who coined the name to include some tetrapods from the Paleozoic that shared some specific characteristics in the notochord and teeth. Lepospondyls have sometimes been considered to be either related or ancestral to modern amphibians or to Amniota. It has been suggested that the grouping is polyphyletic, with aïstopods being primitive stem-tetrapods, while recumbirostran microsaurs are primitive reptiles.
In phylogenetics, long branch attraction (LBA) is a form of systematic error whereby distantly related lineages are incorrectly inferred to be closely related. LBA arises when the amount of molecular or morphological change accumulated within a lineage is sufficient to cause that lineage to appear similar to another long-branched lineage, solely because they have both undergone a large amount of change, rather than because they are related by descent. Such bias is more common when the overall divergence of some taxa results in long branches within a phylogeny. Long branches are often attracted to the base of a phylogenetic tree, because the lineage included to represent an outgroup is often also long-branched. The frequency of true LBA is unclear and often debated, and some authors view it as untestable and therefore irrelevant to empirical phylogenetic inference. Although often viewed as a failing of parsimony-based methodology, LBA could in principle result from a variety of scenarios and be inferred under multiple analytical paradigms.
A grade is a taxon united by a level of morphological or physiological complexity. The term was coined by British biologist Julian Huxley, to contrast with clade, a strictly phylogenetic unit.

Pancrustacea is the clade that comprises all crustaceans, including hexapods. This grouping is contrary to the Atelocerata hypothesis, in which Hexapoda and Myriapoda are sister taxa, and Crustacea are only more distantly related. As of 2010, the Pancrustacea taxon was considered well accepted, with most studies recovering Hexapoda within Crustacea. The clade has also been called Tetraconata, referring to having four cone cells in the ommatidia. This name is preferred by some scientists as a means of avoiding confusion with the use of "pan-" to indicate a clade that includes a crown group and all of its stem group representatives.
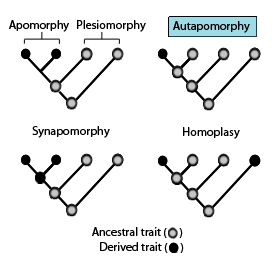
In phylogenetics, an autapomorphy is a distinctive feature, known as a derived trait, that is unique to a given taxon. That is, it is found only in one taxon, but not found in any others or outgroup taxa, not even those most closely related to the focal taxon. It can therefore be considered an apomorphy in relation to a single taxon. The word autapomorphy, introduced in 1950 by German entomologist Willi Hennig, is derived from the Greek words αὐτός, autos "self"; ἀπό, apo "away from"; and μορφή, morphḗ = "shape".
Phylogenetic nomenclature is a method of nomenclature for taxa in biology that uses phylogenetic definitions for taxon names as explained below. This contrasts with the traditional approach, in which taxon names are defined by a type, which can be a specimen or a taxon of lower rank, and a description in words. Phylogenetic nomenclature is currently regulated by the International Code of Phylogenetic Nomenclature (PhyloCode).

Lydekkerinidae is a family of stereospondyl temnospondyls that lived in the Early Triassic period. During this time period, lydekkerinids were widely distributed, with putative remains reported from Russia, Greenland, India, South Africa, Madagascar, Australia, and Antarctica. In contrast to most other stereospondyls, lydekkerinids were relatively small-bodied. The type genus is Lydekkerina, the namesake of the family and the best-known lydekkerinid.

Bigyra is a phylum of microscopic eukaryotes that are found at the base of the Stramenopiles clade. It includes three well-known heterotrophic groups Bicosoecida, Opalinata and Labyrinthulomycetes, as well as several small clades initially discovered through environmental DNA samples: Nanomonadea, Placididea, Opalomonadea and Eogyrea. The classification of Bigyra has changed several times since its origin, and its monophyly remains unresolved.
In bacteriology, a taxon in disguise is a species, genus or higher unit of biological classification whose evolutionary history reveals has evolved from another unit of a similar or lower rank, making the parent unit paraphyletic. That happens when rapid evolution makes a new species appear so radically different from the ancestral group that it is not (initially) recognised as belonging to the parent phylogenetic group, which is left as an evolutionary grade.

Orchideae is a tribe of orchids in the subfamily Orchidoideae. Historically, it was divided into 2 subtribes, Orchidinae and Habenariinae. The subtribe Orchidinae alone contains about 1,800 species. However, although some phylogenetic studies have established the monophyly of the subtribes, the generic boundaries are unclear, with many genera as traditionally circumscribed being paraphyletic or even polyphyletic. Species of genera such as Habenaria and Platanthera have been placed into both subtribes. A 2017 molecular phylogenetic study found that both subtribes did form clades, but did not formally recognize Habenariinae, because of missing genera and uncertainty over generic boundaries. The Asian species of Orchideae, in particular, have been subject to repeated changes of generic placement from 2012 onwards.

Character evolution is the process by which a character or trait evolves along the branches of an evolutionary tree. Character evolution usually refers to single changes within a lineage that make this lineage unique from others. These changes are called character state changes and they are often used in the study of evolution to provide a record of common ancestry. Character state changes can be phenotypic changes, nucleotide substitutions, or amino acid substitutions. These small changes in a species can be identifying features of when exactly a new lineage diverged from an old one.