
Plasmodium is a genus of unicellular eukaryotes that are obligate parasites of vertebrates and insects. The life cycles of Plasmodium species involve development in a blood-feeding insect host which then injects parasites into a vertebrate host during a blood meal. Parasites grow within a vertebrate body tissue before entering the bloodstream to infect red blood cells. The ensuing destruction of host red blood cells can result in malaria. During this infection, some parasites are picked up by a blood-feeding insect, continuing the life cycle.

Coccidia (Coccidiasina) are a subclass of microscopic, spore-forming, single-celled obligate intracellular parasites belonging to the apicomplexan class Conoidasida. As obligate intracellular parasites, they must live and reproduce within an animal cell. Coccidian parasites infect the intestinal tracts of animals, and are the largest group of apicomplexan protozoa.

Eimeria is a genus of apicomplexan parasites that includes various species capable of causing the disease coccidiosis in animals such as cattle, poultry and smaller ruminants including sheep and goats. Eimeria species are considered to be monoxenous because the life cycle is completed within a single host, and stenoxenous because they tend to be host specific, although a number of exceptions have been identified. Species of this genus infect a wide variety of hosts. Thirty-one species are known to occur in bats (Chiroptera), two in turtles, and 130 named species infect fish. Two species infect seals. Five species infect llamas and alpacas: E. alpacae, E. ivitaensis, E. lamae, E. macusaniensis, and E. punonensis. A number of species infect rodents, including E. couesii, E. kinsellai, E. palustris, E. ojastii and E. oryzomysi. Others infect poultry, rabbits and cattle. For full species list, see below.
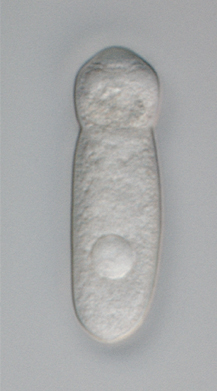
The gregarines are a group of Apicomplexan alveolates, classified as the Gregarinasina or Gregarinia. The large parasites inhabit the intestines of many invertebrates. They are not found in any vertebrates. Gregarines are closely related to both Toxoplasma and Plasmodium, which cause toxoplasmosis and malaria, respectively. Both protists use protein complexes similar to those that are formed by the gregarines for gliding motility and for invading target cells. This makes the gregarines excellent models for studying gliding motility, with the goal of developing treatment options for both toxoplasmosis and malaria. Thousands of different species of gregarine are expected to be found in insects, and 99% of these gregarine species still need to be described. Each insect species can be the host of multiple gregarine species. One of the most-studied gregarines is Gregarina garnhami. In general, gregarines are regarded as a very successful group of parasites, as their hosts are distributed over the entire planet.
An apicoplast is a derived non-photosynthetic plastid found in most Apicomplexa, including Toxoplasma gondii, and Plasmodium falciparum and other Plasmodium spp., but not in others such as Cryptosporidium. It originated from algae through secondary endosymbiosis; there is debate as to whether this was a green or red alga. The apicoplast is surrounded by four membranes within the outermost part of the endomembrane system. The apicoplast hosts important metabolic pathways like fatty acid synthesis, isoprenoid precursor synthesis and parts of the heme biosynthetic pathway.

Adeleorina is a suborder of parasites in the phylum Apicomplexa.
Chromera velia, also known as a "chromerid", is a unicellular photosynthetic organism in the superphylum Alveolata. It is of interest in the study of apicomplexan parasites, specifically their evolution and accordingly, their unique vulnerabilities to drugs.

Myzozoa is a grouping of specific phyla within Alveolata, that either feed through myzocytosis, or were ancestrally capable of feeding through myzocytosis.
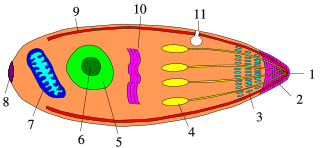
Apicomplexans, a group of intracellular parasites, have life cycle stages that allow them to survive the wide variety of environments they are exposed to during their complex life cycle. Each stage in the life cycle of an apicomplexan organism is typified by a cellular variety with a distinct morphology and biochemistry.
Polychromophilus is a genus of obligate intracellular eukaryotic parasites that infect bats from every continent except Antarctica. They are transmitted by bat flies, which act as an insect vector as well as the parasite’s site of sporogeny. Polychromophilus follows a fairly typical Haemospororidian lifecycle, with gametocytes and gametes restricted to the bloodstream of the host and meronts infecting organs – most notably the lungs and the liver. The type species is Polychromophilus melanipherus, and was described by Dionisi in 1898.
Goussia is a taxonomic genus, first described in 1896 by Labbé, containing parasitic protists which largely target fish and amphibians as their hosts. Members of this genus are homoxenous and often reside in the gastrointestinal tract of the host, however others may be found in organs such as the gallbladder or liver. The genera Goussia, as current phylogenies indicate, is part of the class Conoidasida, which is a subset of the parasitic phylum Apicomplexa; features of this phylum, such as a distinct apical complex containing specialized secretory organelles, an apical polar ring, and a conoid are all present within Goussia, and assist in the mechanical invasion of host tissue. The name Goussia is derived from the French word gousse, meaning pod. This name is based on the bi-valve sporocyst morphology which some Goussians display. Of the original 8 classified Goussians, 6 fit the “pod” morphology. As of this writing, the genera consists of 59 individual species.
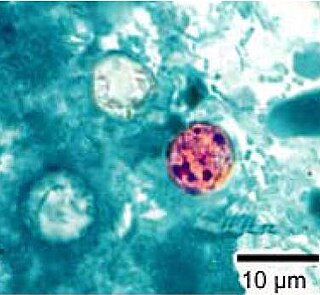
Cyclospora is a genus of apicomplexan parasites. It includes the species Cyclospora cayetanensis, the causative agent of cyclosporiasis. Members of Cyclospora are characterized as having oocysts with two sporocysts, each containing two sporozoites.
The genus Schellackia comprises obligate unicellular eukaryotic parasites within the phylum Apicomplexa, and infects numerous species of lizards and amphibians worldwide. Schellackia is transmitted via insect vectors, primarily mites and mosquitoes, which take up the parasite in blood meals. These vectors then subsequently infect reptilian and amphibian which consume the infected insects. The parasites deform erythrocytes of the host into crescents, and can be visualized using a blood smear.
Vitrella brassicaformis (CCMP3155) is a unicellular alga belonging to the eukaryotic supergroup Alveolata. V. brassicaformis and its closest known relative, Chromera velia, are the only two currently described members of the phylum Chromerida, which in turn constitutes part of the taxonomically unranked group Colpodellida. Chromerida is phylogenetically closely related to the phylum Apicomplexa, which includes Plasmodium, the agent of malaria. Notably, both V. brassicaformis and C. velia are photosynthetic, each containing a complex secondary plastid. This characteristic defined the discovery of these so-called 'chromerids,' as their photosynthetic capacity positioned them to shed light upon the evolution of Apicomplexa's non-photosynthetic parasitism. Both genera lack chlorophyll b or c; these absences link the two taxonomically, as algae bearing only chlorophyll a are rare amid the biodiversity of life. Despite their similarities, V. brassicaformis differs significantly from C. velia in morphology, lifecycle, and accessory photosynthetic pigmentation. V. brassicaformis has a green color, with a complex lifecycle involving multiple pathways and a range of sizes and morphologies, while Chromera has a brown color and cycles through a simpler process from generation to generation. The color differences are due to differences in accessory pigments.
The Archigregarinorida are an order of parasitic alveolates in the phylum Apicomplexa. Species in this order infect marine invertebrates — usually annelids, ascidians, hemichordates and sipunculids.
Lankesterella is a genus in the phylum Apicomplexa. Species in this genus infect amphibians, reptiles and birds.
The Eugregarinorida are the most large and diverse order of gregarines — parasitic protists belonging to the phylum Apicomplexa. Eugregarines are found in marine, freshwater and terrestrial habitats. These species possess large trophozoites that are significantly different in morphology and behavior from the sporozoites. This taxon contains most of the known gregarine species.
Filipodium is a genus of parasites in the phylum Apicomplexa. Species in this genus infect marine invertebrates.
Nematopsis (Nee-mah-top-cis) is a genus gregarine Apicomplexan of the family Porosporidae. It is an aquatic parasite of crustaceans with a molluscan intermediate host. Nematopsis has been distinguished from the similar genus Porospora by its resistant and encapsulated oocyst. Little molecular biology has been performed on the members of the Nemaptosis and species are described based on molluscan and crustacean hosts as well as oocyst structure. A total of 38 species have been described and are found all over the world.

Gregarina garnhami is a eukaryotic unicellular organism belonging to the Apicomplexa described in 1956 by Canning as a parasite found in several locusts, such as the desert locust, African migratory locust, and Egyptian locust. Especially, the desert locust is the host for this species, as up to 100% of animals can become infected. An estimated thousands of different species of gregarines can be in insects and 99% of these gregarines still need to be described. Each insect is said to host multiple species. A remarkable feature of G. garnhami is its autofluorescence.








