
Malaria is a mosquito-borne infectious disease that affects vertebrates. Human malaria causes symptoms that typically include fever, fatigue, vomiting, and headaches. In severe cases, it can cause jaundice, seizures, coma, or death. Symptoms usually begin 10 to 15 days after being bitten by an infected Anopheles mosquito. If not properly treated, people may have recurrences of the disease months later. In those who have recently survived an infection, reinfection usually causes milder symptoms. This partial resistance disappears over months to years if the person has no continuing exposure to malaria.
Antimalarial medications or simply antimalarials are a type of antiparasitic chemical agent, often naturally derived, that can be used to treat or to prevent malaria, in the latter case, most often aiming at two susceptible target groups, young children and pregnant women. As of 2018, modern treatments, including for severe malaria, continued to depend on therapies deriving historically from quinine and artesunate, both parenteral (injectable) drugs, expanding from there into the many classes of available modern drugs. Incidence and distribution of the disease is expected to remain high, globally, for many years to come; moreover, known antimalarial drugs have repeatedly been observed to elicit resistance in the malaria parasite—including for combination therapies featuring artemisinin, a drug of last resort, where resistance has now been observed in Southeast Asia. As such, the needs for new antimalarial agents and new strategies of treatment remain important priorities in tropical medicine. As well, despite very positive outcomes from many modern treatments, serious side effects can impact some individuals taking standard doses.

Plasmodium is a genus of unicellular eukaryotes that are obligate parasites of vertebrates and insects. The life cycles of Plasmodium species involve development in a blood-feeding insect host which then injects parasites into a vertebrate host during a blood meal. Parasites grow within a vertebrate body tissue before entering the bloodstream to infect red blood cells. The ensuing destruction of host red blood cells can result in malaria. During this infection, some parasites are picked up by a blood-feeding insect, continuing the life cycle.

Plasmodium falciparum is a unicellular protozoan parasite of humans, and the deadliest species of Plasmodium that causes malaria in humans. The parasite is transmitted through the bite of a female Anopheles mosquito and causes the disease's most dangerous form, falciparum malaria. It is responsible for around 50% of all malaria cases. P. falciparum is therefore regarded as the deadliest parasite in humans. It is also associated with the development of blood cancer and is classified as a Group 2A (probable) carcinogen.
A trophozoite is the activated, feeding stage in the life cycle of certain protozoa such as malaria-causing Plasmodium falciparum and those of the Giardia group. The complementary form of the trophozoite state is the thick-walled cyst form. They are often different from the cyst stage, which is a protective, dormant form of the protozoa. Trophozoites are often found in the host's body fluids and tissues and in many cases, they are the form of the protozoan that causes disease in the host. In the protozoan, Entamoeba histolytica it invades the intestinal mucosa of its host, causing dysentery, which aid in the trophozoites traveling to the liver and leading to the production of hepatic abscesses.

A gametocyte is a eukaryotic germ cell that divides by mitosis into other gametocytes or by meiosis into gametids during gametogenesis. Male gametocytes are called spermatocytes, and female gametocytes are called oocytes.

Giovanni Battista Grassi was an Italian physician and zoologist, best known for his pioneering works on parasitology, especially on malariology. He was Professor of Comparative Zoology at the University of Catania from 1883, and Professor of Comparative Anatomy at Sapienza University of Rome from 1895 until his death. His first major research on the taxonomy and biology of termites earned him the Royal Society's Darwin Medal in 1896.

Primaquine is a medication used to treat and prevent malaria and to treat Pneumocystis pneumonia. Specifically it is used for malaria due to Plasmodium vivax and Plasmodium ovale along with other medications and for prevention if other options cannot be used. It is an alternative treatment for Pneumocystis pneumonia together with clindamycin. It is taken by mouth.
Recrudescence is the recurrence of an undesirable condition. In medicine, it is usually defined as the recurrence of symptoms after a period of remission or quiescence, in which sense it can sometimes be synonymous with relapse. In a narrower sense, it can also be such a recurrence with higher severity than before the remission. "Relapse" conventionally has a specific meaning when used in relation to malaria.

Plasmodium ovale is a species of parasitic protozoon that causes tertian malaria in humans. It is one of several species of Plasmodium parasites that infect humans, including Plasmodium falciparum and Plasmodium vivax which are responsible for most cases of malaria in the world. P. ovale is rare compared to these two parasites, and substantially less dangerous than P. falciparum.

Plasmodium malariae is a parasitic protozoan that causes malaria in humans. It is one of several species of Plasmodium parasites that infect other organisms as pathogens, also including Plasmodium falciparum and Plasmodium vivax, responsible for most malarial infection. Found worldwide, it causes a so-called "benign malaria", not nearly as dangerous as that produced by P. falciparum or P. vivax. The signs include fevers that recur at approximately three-day intervals – a quartan fever or quartan malaria – longer than the two-day (tertian) intervals of the other malarial parasite.

Plasmodium knowlesi is a parasite that causes malaria in humans and other primates. It is found throughout Southeast Asia, and is the most common cause of human malaria in Malaysia. Like other Plasmodium species, P. knowlesi has a life cycle that requires infection of both a mosquito and a warm-blooded host. While the natural warm-blooded hosts of P. knowlesi are likely various Old World monkeys, humans can be infected by P. knowlesi if they are fed upon by infected mosquitoes. P. knowlesi is a eukaryote in the phylum Apicomplexa, genus Plasmodium, and subgenus Plasmodium. It is most closely related to the human parasite Plasmodium vivax as well as other Plasmodium species that infect non-human primates.

Tafenoquine, sold under the brand name Krintafel among others, is a medication used to prevent and to treat malaria. With respect to acute malaria, it is used together with other medications to prevent relapse by Plasmodium vivax. It may be used to prevent all types of malaria. It is taken by mouth.

The history of malaria extends from its prehistoric origin as a zoonotic disease in the primates of Africa through to the 21st century. A widespread and potentially lethal human infectious disease, at its peak malaria infested every continent except Antarctica. Its prevention and treatment have been targeted in science and medicine for hundreds of years. Since the discovery of the Plasmodium parasites which cause it, research attention has focused on their biology as well as that of the mosquitoes which transmit the parasites.
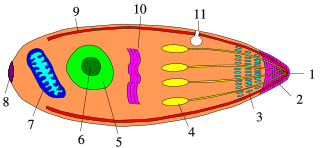
Apicomplexans, a group of intracellular parasites, have life cycle stages that allow them to survive the wide variety of environments they are exposed to during their complex life cycle. Each stage in the life cycle of an apicomplexan organism is typified by a cellular variety with a distinct morphology and biochemistry.
Polychromophilus is a genus of obligate intracellular eukaryotic parasites that infect bats from every continent except Antarctica. They are transmitted by bat flies, which act as an insect vector as well as the parasite’s site of sporogeny. Polychromophilus follows a fairly typical Haemospororidian lifecycle, with gametocytes and gametes restricted to the bloodstream of the host and meronts infecting organs – most notably the lungs and the liver. The type species is Polychromophilus melanipherus, and was described by Dionisi in 1898.
Plasmodium coatneyi is a parasitic species that is an agent of malaria in nonhuman primates. P. coatneyi occurs in Southeast Asia. The natural host of this species is the rhesus macaque and crab-eating macaque, but there has been no evidence that zoonosis of P. coatneyi can occur through its vector, the female Anopheles mosquito.
Circumsporozoite protein (CSP) is a secreted protein of the sporozoite stage of the malaria parasite and is the antigenic target of RTS,S and other malaria vaccines. The amino-acid sequence of CSP consists of an immunodominant central repeat region flanked by conserved motifs at the N- and C- termini that are implicated in protein processing as the parasite travels from the mosquito to the mammalian vector. The amino acid sequence of CSP was determined in 1984.

Plasmodium cynomolgi is an apicomplexan parasite that infects mosquitoes and Asian Old World monkeys. In recent years, a number of natural infections of humans have also been documented. This species has been used as a model for human Plasmodium vivax because Plasmodium cynomolgi shares the same life cycle and some important biological features with P. vivax.

Quartan fever is one of the four types of malaria which can be contracted by humans.


