
Banded iron formations are distinctive units of sedimentary rock consisting of alternating layers of iron oxides and iron-poor chert. They can be up to several hundred meters in thickness and extend laterally for several hundred kilometers. Almost all of these formations are of Precambrian age and are thought to record the oxygenation of the Earth's oceans. Some of the Earth's oldest rock formations, which formed about 3,700 million years ago (Ma), are associated with banded iron formations.

Stromatolites or stromatoliths are layered sedimentary formations (microbialite) that are created mainly by photosynthetic microorganisms such as cyanobacteria, sulfate-reducing bacteria, and Pseudomonadota. These microorganisms produce adhesive compounds that cement sand and other rocky materials to form mineral "microbial mats". In turn, these mats build up layer by layer, growing gradually over time. A stromatolite may grow to a meter or more. Fossilized stromatolites provide important records of some of the most ancient life. As of the Holocene, living forms are rare.

Chert is a hard, fine-grained sedimentary rock composed of microcrystalline or cryptocrystalline quartz, the mineral form of silicon dioxide (SiO2). Chert is characteristically of biological origin, but may also occur inorganically as a chemical precipitate or a diagenetic replacement, as in petrified wood.

A concretion is a hard, compact mass formed by the precipitation of mineral cement within the spaces between particles, and is found in sedimentary rock or soil. Concretions are often ovoid or spherical in shape, although irregular shapes also occur. The word 'concretion' is derived from the Latin concretio "(act of) compacting, condensing, congealing, uniting", itself from con meaning 'together' and crescere meaning "to grow". Concretions form within layers of sedimentary strata that have already been deposited. They usually form early in the burial history of the sediment, before the rest of the sediment is hardened into rock. This concretionary cement often makes the concretion harder and more resistant to weathering than the host stratum.

The Gunflint Range is an iron ore deposit in northern Minnesota in the United States and Northwestern Ontario, Canada. The range extends from the extreme northern portion of Cook County, Minnesota into the Thunder Bay District, Ontario.
The Cuyuna Range is an inactive iron range to the southwest of the Mesabi Range, largely within Crow Wing County, Minnesota. It lies along a 68-mile-long (109 km) line between Brainerd, Minnesota, and Aitkin, Minnesota. The width ranges from 1 to 10 miles.
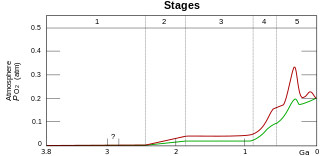
The Great Oxidation Event (GOE) or Great Oxygenation Event, also called the Oxygen Catastrophe, Oxygen Revolution, Oxygen Crisis or Oxygen Holocaust, was a time interval during the Early Earth's Paleoproterozoic Era when the Earth's atmosphere and the shallow ocean first experienced a rise in the concentration of oxygen. This began approximately 2.460–2.426 Ga (billion years) ago, during the Siderian period, and ended approximately 2.060 Ga, during the Rhyacian. Geological, isotopic, and chemical evidence suggests that biologically-produced molecular oxygen (dioxygen or O2) started to accumulate in Earth's atmosphere and changed it from a weakly reducing atmosphere practically devoid of oxygen into an oxidizing one containing abundant free oxygen, with oxygen levels being as high as 10% of their present atmospheric level by the end of the GOE.

The Pilbara Craton is an old and stable part of the continental lithosphere located in the Pilbara region of Western Australia.
Elso Sterrenberg Barghoorn was an American paleobotanist, called by his student Andrew Knoll, the present Fisher Professor of Natural History at Harvard, "the father of Pre-Cambrian palaeontology."

Bitter Springs Group is a Precambrian fossil locality in Australia, which preserves microorganisms in silica. Its preservational mode ceased in the late Precambrian with the advent of silicifying organisms.

Permineralization is a process of fossilization of bones and tissues in which mineral deposits form internal casts of organisms. Carried by water, these minerals fill the spaces within organic tissue. Because of the nature of the casts, permineralization is particularly useful in studies of the internal structures of organisms, usually of plants.
Kakabekia is a genus of microorganism. Kakabekia umbellata was first found in the 1.88 billion year old Gunflint Chert, and in 1966, a living member of the genus, Kakabekia barghoorniana was discovered in Wales - it is also found around the world, typically at 1000-2000m elevations. The roughly 2 billion year interval between these species has led some to call it the “oldest living fossil”.

The Animikie Group is a geologic group composed of sedimentary and metasedimentary rock, having been originally deposited between 2,500 and 1,800 million years ago during the Paleoproterozoic era, within the Animikie Basin. This group of formations is geographically divided into the Gunflint Range, the Mesabi and Vermilion ranges, and the Cuyuna Range. On the map, the Animikie Group is the dark gray northeast-trending belt which ranges from south-central Minnesota, U.S., up to Thunder Bay, Ontario, Canada. The Gunflint Iron Range is the linear black formation labeled G, the Mesabi Iron Range is the jagged black linear formation labeled F, and Cuyuna Iron Range is the two black spots labeled E. The gabbro of the Duluth Complex, intruded during the formation of the Midcontinent Rift, separates the Mesabi and Gunflint iron ranges; it is shown by the speckled area wrapping around the western end of Lake Superior.

Iron-rich sedimentary rocks are sedimentary rocks which contain 15 wt.% or more iron. However, most sedimentary rocks contain iron in varying degrees. The majority of these rocks were deposited during specific geologic time periods: The Precambrian, the early Paleozoic, and the middle to late Mesozoic. Overall, they make up a very small portion of the total sedimentary record.

The Barberton Greenstone Belt of eastern South Africa contains some of the most widely accepted fossil evidence for Archean life. These cell-sized prokaryote fossils are seen in the Barberton fossil record in rocks as old as 3.5 billion years. The Barberton Greenstone Belt is an excellent place to study the Archean Earth due to exposed sedimentary and metasedimentary rocks.
James William Schopf is an American paleobiologist and professor of earth sciences at the University of California Los Angeles. He is also Director of the Center for the Study of Evolution and the Origin of Life, and a member of the Department of Earth and Space Sciences, the Institute of Geophysics and Planetary Physics, and the Molecular Biology Institute at UCLA. He is most well known for his study of Precambrian prokaryotic life in Australia's Apex chert. Schopf has published extensively in the peer reviewed literature about the origins of life on Earth. He is the first to discover Precambrian microfossils in stromatolitic sediments of Australia (1965), South Africa (1966), Russia (1977), India (1978), and China (1984). He served as NASA's principal investigator of lunar samples during 1969–1974.
Stanley Awramik is an American biogeologist and paleontologist. He is best known for his work related to the Precambrian. In 2013, he was inducted as a fellow of the Geological Society of America.

Microbialite is a benthic sedimentary deposit made of carbonate mud that is formed with the mediation of microbes. The constituent carbonate mud is a type of automicrite ; therefore, it precipitates in situ instead of being transported and deposited. Being formed in situ, a microbialite can be seen as a type of boundstone where reef builders are microbes, and precipitation of carbonate is biotically induced instead of forming tests, shells or skeletons.
The Dresser Formation is a Paleoarchean geologic formation that outcrops as a generally circular ring of hills the North Pole Dome area of the East Pilbara Terrane of the Pilbara Craton of Western Australia. This formation is one of many formations that comprise the Warrawoona Group, which is the lowermost of four groups that comprise the Pilbara Supergroup. The Dresser Formation is part of the Panorama greenstone belt that surrounds and outcrops around the intrusive North Pole Monzogranite. Dresser Formation consists of metamorphosed, blue, black, and white bedded chert; pillow basalt; carbonate rocks; minor felsic volcaniclastic sandstone and conglomerate; hydrothermal barite; evaporites; and stromatolites. The lowermost of three stratigraphic units that comprise the Dresser Formation contains some of the Earth's earliest commonly accepted evidence of life such as morphologically diverse stromatolites, microbially induced sedimentary structures, putative organic microfossils, and biologically fractionated carbon and sulfur isotopic data.
The Allamoore Formation is a Precambrian geologic formation found in the vicinity of Van Horn, Texas. The formation is notable for the well-preserved microfossils found in chert beds within the formation. These may include early eukaryotes. Possible fossil burrows were reported in the formation in 1995, which would push the origin of the first animals (Metazoa) back to before 1000 million years ago. However, these have since been explained as diagenetic structures, formed by nonbiological processes as the sediments making up the formation were compacted and lithified.
