
Leptospermum scoparium, commonly called mānuka, mānuka myrtle, New Zealand teatree, broom tea-tree, or just tea tree, is a species of flowering plant in the myrtle family Myrtaceae, native to New Zealand and south-east Australia. Its nectar produces Mānuka honey.

The human microbiome is the aggregate of all microbiota that reside on or within human tissues and biofluids along with the corresponding anatomical sites in which they reside, including the skin, mammary glands, seminal fluid, uterus, ovarian follicles, lung, saliva, oral mucosa, conjunctiva, biliary tract, and gastrointestinal tract. Types of human microbiota include bacteria, archaea, fungi, protists, and viruses. Though micro-animals can also live on the human body, they are typically excluded from this definition. In the context of genomics, the term human microbiome is sometimes used to refer to the collective genomes of resident microorganisms; however, the term human metagenome has the same meaning.

Aeroplankton are tiny lifeforms that float and drift in the air, carried by wind. Most of the living things that make up aeroplankton are very small to microscopic in size, and many can be difficult to identify because of their tiny size. Scientists collect them for study in traps and sweep nets from aircraft, kites or balloons. The study of the dispersion of these particles is called aerobiology.

Gut microbiota, gut microbiome, or gut flora, are the microorganisms, including bacteria, archaea, fungi, and viruses, that live in the digestive tracts of animals. The gastrointestinal metagenome is the aggregate of all the genomes of the gut microbiota. The gut is the main location of the human microbiome. The gut microbiota has broad impacts, including effects on colonization, resistance to pathogens, maintaining the intestinal epithelium, metabolizing dietary and pharmaceutical compounds, controlling immune function, and even behavior through the gut–brain axis.
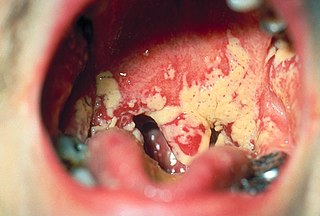
Oral microbiology is the study of the microorganisms (microbiota) of the oral cavity and their interactions between oral microorganisms or with the host. The environment present in the human mouth is suited to the growth of characteristic microorganisms found there. It provides a source of water and nutrients, as well as a moderate temperature. Resident microbes of the mouth adhere to the teeth and gums to resist mechanical flushing from the mouth to stomach where acid-sensitive microbes are destroyed by hydrochloric acid.

Microbiota are the range of microorganisms that may be commensal, mutualistic, or pathogenic found in and on all multicellular organisms, including plants. Microbiota include bacteria, archaea, protists, fungi, and viruses, and have been found to be crucial for immunologic, hormonal, and metabolic homeostasis of their host.

Oral ecology is the microbial ecology of the microorganisms found in mouths. Oral ecology, like all forms of ecology, involves the study of the living things found in oral cavities as well as their interactions with each other and with their environment. Oral ecology is frequently investigated from the perspective of oral disease prevention, often focusing on conditions such as dental caries, candidiasis ("thrush"), gingivitis, periodontal disease, and others. However, many of the interactions between the microbiota and oral environment protect from disease and support a healthy oral cavity. Interactions between microbes and their environment can result in the stabilization or destabilization of the oral microbiome, with destabilization believed to result in disease states. Destabilization of the microbiome can be influenced by several factors, including diet changes, drugs or immune system disorders.

Mānuka honey is a monofloral honey produced from the nectar of the mānuka tree, Leptospermum scoparium.
Soil microbiology is the study of microorganisms in soil, their functions, and how they affect soil properties. It is believed that between two and four billion years ago, the first ancient bacteria and microorganisms came about on Earth's oceans. These bacteria could fix nitrogen, in time multiplied, and as a result released oxygen into the atmosphere. This led to more advanced microorganisms, which are important because they affect soil structure and fertility. Soil microorganisms can be classified as bacteria, actinomycetes, fungi, algae and protozoa. Each of these groups has characteristics that define them and their functions in soil.
Microbial biogeography is a subset of biogeography, a field that concerns the distribution of organisms across space and time. Although biogeography traditionally focused on plants and larger animals, recent studies have broadened this field to include distribution patterns of microorganisms. This extension of biogeography to smaller scales—known as "microbial biogeography"—is enabled by ongoing advances in genetic technologies.

The root microbiome is the dynamic community of microorganisms associated with plant roots. Because they are rich in a variety of carbon compounds, plant roots provide unique environments for a diverse assemblage of soil microorganisms, including bacteria, fungi and archaea. The microbial communities inside the root and in the rhizosphere are distinct from each other, and from the microbial communities of bulk soil, although there is some overlap in species composition.
The phycosphere is a microscale mucus region that is rich in organic matter surrounding a phytoplankton cell. This area is high in nutrients due to extracellular waste from the phytoplankton cell and it has been suggested that bacteria inhabit this area to feed on these nutrients. This high nutrient environment creates a microbiome and a diverse food web for microbes such as bacteria and protists. It has also been suggested that the bacterial assemblages within the phycosphere are species-specific and can vary depending on different environmental factors.
A microbiome is the community of microorganisms that can usually be found living together in any given habitat. It was defined more precisely in 1988 by Whipps et al. as "a characteristic microbial community occupying a reasonably well-defined habitat which has distinct physio-chemical properties. The term thus not only refers to the microorganisms involved but also encompasses their theatre of activity". In 2020, an international panel of experts published the outcome of their discussions on the definition of the microbiome. They proposed a definition of the microbiome based on a revival of the "compact, clear, and comprehensive description of the term" as originally provided by Whipps et al., but supplemented with two explanatory paragraphs. The first explanatory paragraph pronounces the dynamic character of the microbiome, and the second explanatory paragraph clearly separates the term microbiota from the term microbiome.
The initial acquisition of microbiota is the formation of an organism's microbiota immediately before and after birth. The microbiota are all the microorganisms including bacteria, archaea and fungi that colonize the organism. The microbiome is another term for microbiota or can refer to the collected genomes.
A holobiont is an assemblage of a host and the many other species living in or around it, which together form a discrete ecological unit through symbiosis, though there is controversy over this discreteness. The components of a holobiont are individual species or bionts, while the combined genome of all bionts is the hologenome. The holobiont concept was initially introduced by the German theoretical biologist Adolf Meyer-Abich in 1943, and then apparently independently by Dr. Lynn Margulis in her 1991 book Symbiosis as a Source of Evolutionary Innovation. The concept has evolved since the original formulations. Holobionts include the host, virome, microbiome, and any other organisms which contribute in some way to the functioning of the whole. Well-studied holobionts include reef-building corals and humans.

All animals on Earth form associations with microorganisms, including protists, bacteria, archaea, fungi, and viruses. In the ocean, animal–microbial relationships were historically explored in single host–symbiont systems. However, new explorations into the diversity of marine microorganisms associating with diverse marine animal hosts is moving the field into studies that address interactions between the animal host and a more multi-member microbiome. The potential for microbiomes to influence the health, physiology, behavior, and ecology of marine animals could alter current understandings of how marine animals adapt to change, and especially the growing climate-related and anthropogenic-induced changes already impacting the ocean environment.
The plant microbiome, also known as the phytomicrobiome, plays roles in plant health and productivity and has received significant attention in recent years. The microbiome has been defined as "a characteristic microbial community occupying a reasonably well-defined habitat which has distinct physio-chemical properties. The term thus not only refers to the microorganisms involved but also encompasses their theatre of activity".
Since the colonization of land by ancestral plant lineages 450 million years ago, plants and their associated microbes have been interacting with each other, forming an assemblage of species that is often referred to as a holobiont. Selective pressure acting on holobiont components has likely shaped plant-associated microbial communities and selected for host-adapted microorganisms that impact plant fitness. However, the high microbial densities detected on plant tissues, together with the fast generation time of microbes and their more ancient origin compared to their host, suggest that microbe-microbe interactions are also important selective forces sculpting complex microbial assemblages in the phyllosphere, rhizosphere, and plant endosphere compartments.
Some microorganisms, such as endophytes, penetrate and occupy the plant internal tissues, forming the endospheric microbiome. The arbuscular mycorrhizal and other endophytic fungi are the dominant colonizers of the endosphere. Bacteria, and to some degree archaea, are important members of endosphere communities. Some of these endophytic microbes interact with their host and provide obvious benefits to plants. Unlike the rhizosphere and the rhizoplane, the endospheres harbor highly specific microbial communities. The root endophytic community can be very distinct from that of the adjacent soil community. In general, diversity of the endophytic community is lower than the diversity of the microbial community outside the plant. The identity and diversity of the endophytic microbiome of above-and below-ground tissues may also differ within the plant.
The cetacean microbiome is the group of communities of microorganisms that reside within whales.







