
The thalamus is a large mass of gray matter located in the dorsal part of the diencephalon. Nerve fibers project out of the thalamus to the cerebral cortex in all directions, known as the thalamocortical radiations, allowing hub-like exchanges of information. It has several functions, such as the relaying of sensory and motor signals to the cerebral cortex and the regulation of consciousness, sleep, and alertness.
The development of the nervous system, or neural development (neurodevelopment), refers to the processes that generate, shape, and reshape the nervous system of animals, from the earliest stages of embryonic development to adulthood. The field of neural development draws on both neuroscience and developmental biology to describe and provide insight into the cellular and molecular mechanisms by which complex nervous systems develop, from nematodes and fruit flies to mammals.

Sonic hedgehog protein (SHH) is encoded for by the SHH gene. The protein is named after the character Sonic the Hedgehog.

In zoology and developmental anatomy, the notochord is an elastic, rod-like anatomical structure found in many deuterostomal animals. A notochord is one of five synapomorphies used to define a species as a chordate.

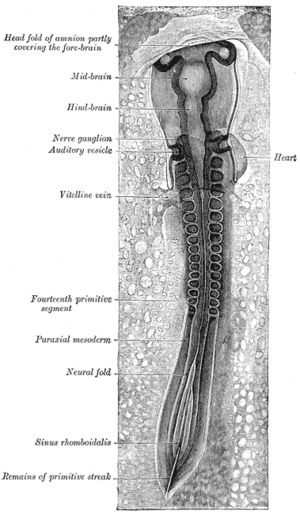
Neurulation refers to the folding process in vertebrate embryos, which includes the transformation of the neural plate into the neural tube. The embryo at this stage is termed the neurula.
In the vertebrate embryo, a rhombomere is a transiently divided segment of the developing neural tube, within the hindbrain region in the area that will eventually become the rhombencephalon. The rhombomeres appear as a series of slightly constricted swellings in the neural tube, caudal to the cephalic flexure. In human embryonic development, the rhombomeres are present by day 29.

Neuromeres are distinct groups of neural crest cells, forming segments in the neural tube of the early embryonic development of the brain. There are three classes of neuromeres in the central nervous system – prosomeres, mesomeres and rhombomeres that will develop the forebrain, midbrain, and hindbrain respectively.

The neural plate is a key developmental structure that serves as the basis for the nervous system. Cranial to the primitive node of the embryonic primitive streak, ectodermal tissue thickens and flattens to become the neural plate. The region anterior to the primitive node can be generally referred to as the neural plate. Cells take on a columnar appearance in the process as they continue to lengthen and narrow. The ends of the neural plate, known as the neural folds, push the ends of the plate up and together, folding into the neural tube, a structure critical to brain and spinal cord development. This process as a whole is termed primary neurulation.
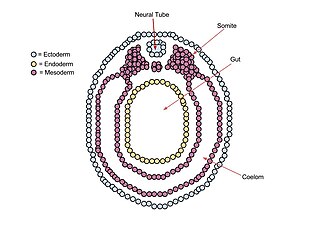
A neurula is a vertebrate embryo at the early stage of development in which neurulation occurs. The neurula stage is preceded by the gastrula stage; consequentially, neurulation is preceded by gastrulation. Neurulation marks the beginning of the process of organogenesis.
Neural crest cells are a temporary group of cells that arise from the embryonic ectoderm germ layer, and in turn give rise to a diverse cell lineage—including melanocytes, craniofacial cartilage and bone, smooth muscle, peripheral and enteric neurons and glia.
Zinc finger protein GLI1 also known as glioma-associated oncogene is a protein that in humans is encoded by the GLI1 gene. It was originally isolated from human glioblastoma cells.

The neural fold is a structure that arises during neurulation in the embryonic development of both birds and mammals among other organisms. This structure is associated with primary neurulation, meaning that it forms by the coming together of tissue layers, rather than a clustering, and subsequent hollowing out, of individual cells. In humans, the neural folds are responsible for the formation of the anterior end of the neural tube. The neural folds are derived from the neural plate, a preliminary structure consisting of elongated ectoderm cells. The folds give rise to neural crest cells, as well as bringing about the formation of the neural tube.
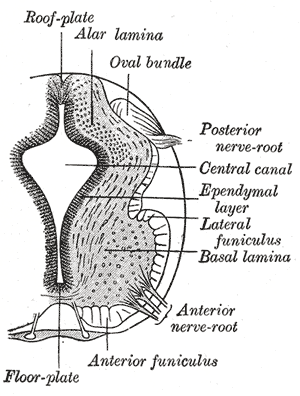
The floor plate is a structure integral to the developing nervous system of vertebrate organisms. Located on the ventral midline of the embryonic neural tube, the floor plate is a specialized glial structure that spans the anteroposterior axis from the midbrain to the tail regions. It has been shown that the floor plate is conserved among vertebrates, such as zebrafish and mice, with homologous structures in invertebrates such as the fruit fly Drosophila and the nematode C. elegans. Functionally, the structure serves as an organizer to ventralize tissues in the embryo as well as to guide neuronal positioning and differentiation along the dorsoventral axis of the neural tube.

The zona limitans intrathalamica (ZLI) is a lineage-restriction compartment and primary developmental boundary in the vertebrate forebrain that serves as a signaling center and a restrictive border between the thalamus and the prethalamus.

Limb development in vertebrates is an area of active research in both developmental and evolutionary biology, with much of the latter work focused on the transition from fin to limb.

The development of fishes is unique in some specific aspects compared to the development of other animals.

Ras-related protein Rab-23 is a protein that in humans is encoded by the RAB23 gene. Alternative splicing occurs at this gene locus and two transcript variants encoding the same protein have been identified.
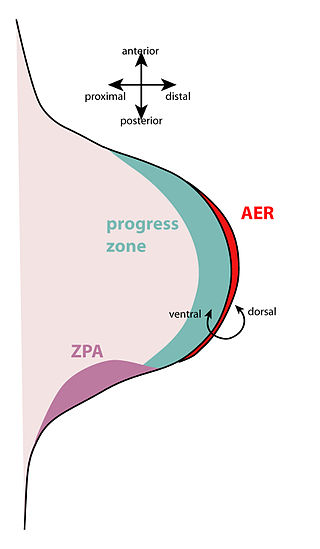
The zone of polarizing activity (ZPA) is an area of mesenchyme that contains signals which instruct the developing limb bud to form along the anterior/posterior axis. Limb bud is undifferentiated mesenchyme enclosed by an ectoderm covering. Eventually, the limb bud develops into bones, tendons, muscles and joints. Limb bud development relies not only on the ZPA, but also many different genes, signals, and a unique region of ectoderm called the apical ectodermal ridge (AER). Research by Saunders and Gasseling in 1948 identified the AER and its subsequent involvement in proximal distal outgrowth. Twenty years later, the same group did transplantation studies in chick limb bud and identified the ZPA. It wasn't until 1993 that Todt and Fallon showed that the AER and ZPA are dependent on each other.
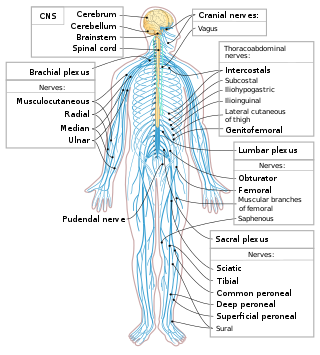
The spinal cord is a long, thin, tubular structure made up of nervous tissue that extends from the medulla oblongata in the brainstem to the lumbar region of the vertebral column (backbone) of vertebrate animals. The center of the spinal cord is hollow and contains a structure called central canal, which contains cerebrospinal fluid. The spinal cord is also covered by meninges and enclosed by the neural arches. Together, the brain and spinal cord make up the central nervous system.
The development of the cerebral cortex, known as corticogenesis is the process during which the cerebral cortex of the brain is formed as part of the development of the nervous system of mammals including its development in humans. The cortex is the outer layer of the brain and is composed of up to six layers. Neurons formed in the ventricular zone migrate to their final locations in one of the six layers of the cortex. The process occurs from embryonic day 10 to 17 in mice and between gestational weeks seven to 18 in humans.




{kind=link}