Ontogeny is the origination and development of an organism, usually from the time of fertilization of the egg to adult. The term can also be used to refer to the study of the entirety of an organism's lifespan.

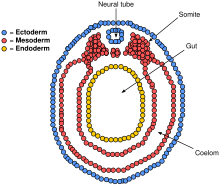
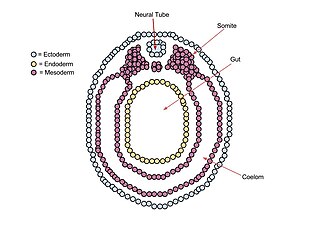
The mesoderm is the middle layer of the three germ layers that develops during gastrulation in the very early development of the embryo of most animals. The outer layer is the ectoderm, and the inner layer is the endoderm.
The development of the nervous system, or neural development (neurodevelopment), refers to the processes that generate, shape, and reshape the nervous system of animals, from the earliest stages of embryonic development to adulthood. The field of neural development draws on both neuroscience and developmental biology to describe and provide insight into the cellular and molecular mechanisms by which complex nervous systems develop, from nematodes and fruit flies to mammals.

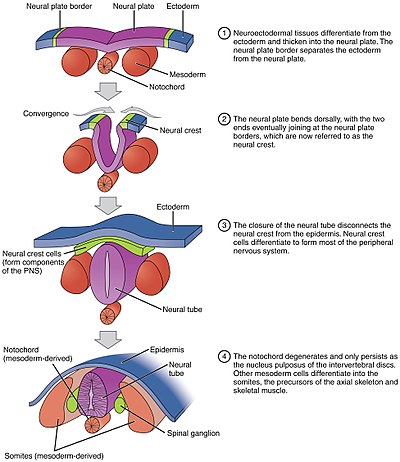
In the developing chordate, the neural tube is the embryonic precursor to the central nervous system, which is made up of the brain and spinal cord. The neural groove gradually deepens as the neural fold become elevated, and ultimately the folds meet and coalesce in the middle line and convert the groove into the closed neural tube. In humans, neural tube closure usually occurs by the fourth week of pregnancy.

The ectoderm is one of the three primary germ layers formed in early embryonic development. It is the outermost layer, and is superficial to the mesoderm and endoderm. It emerges and originates from the outer layer of germ cells. The word ectoderm comes from the Greek ektos meaning "outside", and derma meaning "skin".

Sonic hedgehog protein (SHH) is encoded for by the SHH gene. The protein is named after the character Sonic the Hedgehog.
A germ layer is a primary layer of cells that forms during embryonic development. The three germ layers in vertebrates are particularly pronounced; however, all eumetazoans produce two or three primary germ layers. Some animals, like cnidarians, produce two germ layers making them diploblastic. Other animals such as bilaterians produce a third layer between these two layers, making them triploblastic. Germ layers eventually give rise to all of an animal's tissues and organs through the process of organogenesis.
Organogenesis is the phase of embryonic development that starts at the end of gastrulation and continues until birth. During organogenesis, the three germ layers formed from gastrulation form the internal organs of the organism.

In developmental biology, animal embryonic development, also known as animal embryogenesis, is the developmental stage of an animal embryo. Embryonic development starts with the fertilization of an egg cell (ovum) by a sperm cell, (spermatozoon). Once fertilized, the ovum becomes a single diploid cell known as a zygote. The zygote undergoes mitotic divisions with no significant growth and cellular differentiation, leading to development of a multicellular embryo after passing through an organizational checkpoint during mid-embryogenesis. In mammals, the term refers chiefly to the early stages of prenatal development, whereas the terms fetus and fetal development describe later stages.
The neural plate is a key developmental structure that serves as the basis for the nervous system. Cranial to the primitive node of the embryonic primitive streak, ectodermal tissue thickens and flattens to become the neural plate. The region anterior to the primitive node can be generally referred to as the neural plate. Cells take on a columnar appearance in the process as they continue to lengthen and narrow. The ends of the neural plate, known as the neural folds, push the ends of the plate up and together, folding into the neural tube, a structure critical to brain and spinal cord development. This process as a whole is termed primary neurulation.
A neurula is a vertebrate embryo at the early stage of development in which neurulation occurs. The neurula stage is preceded by the gastrula stage; consequentially, neurulation is preceded by gastrulation. Neurulation marks the beginning of the process of organogenesis.
The primitive node is the organizer for gastrulation in most amniote embryos. In birds it is known as Hensen's node, and in amphibians it is known as the Spemann-Mangold organizer. It is induced by the Nieuwkoop center in amphibians, or by the posterior marginal zone in amniotes including birds.

The neural fold is a structure that arises during neurulation in the embryonic development of both birds and mammals among other organisms. This structure is associated with primary neurulation, meaning that it forms by the coming together of tissue layers, rather than a clustering, and subsequent hollowing out, of individual cells. In humans, the neural folds are responsible for the formation of the anterior end of the neural tube. The neural folds are derived from the neural plate, a preliminary structure consisting of elongated ectoderm cells. The folds give rise to neural crest cells, as well as bringing about the formation of the neural tube.
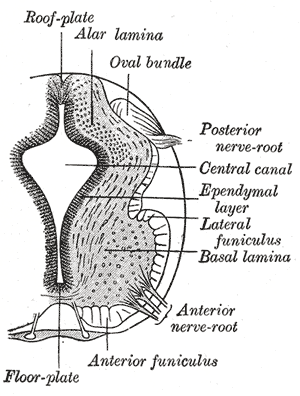
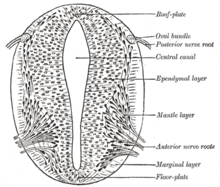
The floor plate is a structure integral to the developing nervous system of vertebrate organisms. Located on the ventral midline of the embryonic neural tube, the floor plate is a specialized glial structure that spans the anteroposterior axis from the midbrain to the tail regions. It has been shown that the floor plate is conserved among vertebrates, such as zebrafish and mice, with homologous structures in invertebrates such as the fruit fly Drosophila and the nematode C. elegans. Functionally, the structure serves as an organizer to ventralize tissues in the embryo as well as to guide neuronal positioning and differentiation along the dorsoventral axis of the neural tube.
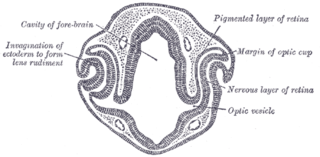
Eye formation in the human embryo begins at approximately three weeks into embryonic development and continues through the tenth week. Cells from both the mesodermal and the ectodermal tissues contribute to the formation of the eye. Specifically, the eye is derived from the neuroepithelium, surface ectoderm, and the extracellular mesenchyme which consists of both the neural crest and mesoderm.
Human embryonic development or human embryogenesis is the development and formation of the human embryo. It is characterised by the processes of cell division and cellular differentiation of the embryo that occurs during the early stages of development. In biological terms, the development of the human body entails growth from a one-celled zygote to an adult human being. Fertilization occurs when the sperm cell successfully enters and fuses with an egg cell (ovum). The genetic material of the sperm and egg then combine to form the single cell zygote and the germinal stage of development commences. Embryonic development in the human, covers the first eight weeks of development; at the beginning of the ninth week the embryo is termed a fetus. The eight weeks has 23 stages.

Johannes Holtfreter was a German-American developmental biologist whose primary focus was the “organizer,” a part of the embryo essential for the development of the proper body plan.
The rostral neuropore or anterior neuropore is a region corresponding to the opening of the embryonic neural tube in the anterior portion of the developing prosencephalon. The central nervous system develops from the neural tube, which initially starts as a plate of cells in the ectoderm and this is called the neural plate, the neural plate then undergoes folding and starts closing from the center of the developing fetus, this leads to two open ends, one situated cranially/rostrally and the other caudally. Bending of the neural plate begins on day 22, and the cranial neuropore closes on day 24. giving rise to the lamina terminalis of the brain.
The Spemann-Mangold organizer is a group of cells that are responsible for the induction of the neural tissues during development in amphibian embryos. First described in 1924 by Hans Spemann and Hilde Mangold, the introduction of the organizer provided evidence that the fate of cells can be influenced by factors from other cell populations. This discovery significantly impacted the world of developmental biology and fundamentally changed the understanding of early development.
A developmental signaling center is defined as a group of cells that release various morphogens which can determine the fates, or destined cell types, of adjacent cells. This process in turn determines what tissues the adjacent cells will form. Throughout the years, various development signaling centers have been discovered.