
Silicon is a chemical element; it has symbol Si and atomic number 14. It is a hard, brittle crystalline solid with a blue-grey metallic luster, and is a tetravalent metalloid and semiconductor. It is a member of group 14 in the periodic table: carbon is above it; and germanium, tin, lead, and flerovium are below it. It is relatively unreactive. Silicon (Si) element is a significant element that is essential for several physiological and metabolic processes in plants. Si is widely regarded as the predominant semiconductor material due to its versatile applications in various electrical devices such as transistors, solar cells, integrated circuits, and others. These may be due to its significant band gap, expansive optical transmission range, extensive absorption spectrum, surface roughening, and effective anti-reflection coating.

A diatom is any member of a large group comprising several genera of algae, specifically microalgae, found in the oceans, waterways and soils of the world. Living diatoms make up a significant portion of the Earth's biomass: they generate about 20 to 50 percent of the oxygen produced on the planet each year, take in over 6.7 billion tonnes of silicon each year from the waters in which they live, and constitute nearly half of the organic material found in the oceans. The shells of dead diatoms can reach as much as a half-mile deep on the ocean floor, and the entire Amazon basin is fertilized annually by 27 million tons of diatom shell dust transported by transatlantic winds from the African Sahara, much of it from the Bodélé Depression, which was once made up of a system of fresh-water lakes.

Biomineralization, also written biomineralisation, is the process by which living organisms produce minerals, often resulting in hardened or stiffened mineralized tissues. It is an extremely widespread phenomenon: all six taxonomic kingdoms contain members that are able to form minerals, and over 60 different minerals have been identified in organisms. Examples include silicates in algae and diatoms, carbonates in invertebrates, and calcium phosphates and carbonates in vertebrates. These minerals often form structural features such as sea shells and the bone in mammals and birds.
In taxonomy, Stappia is a genus of the Hyphomicrobiales. Some members of the genus oxidize carbon monoxide (CO) aerobically. Stappia indica is a diatom associated bacterium which is known to inhibit the growth of diatoms such as Thalassiosira pseudonana.

Siliceous ooze is a type of biogenic pelagic sediment located on the deep ocean floor. Siliceous oozes are the least common of the deep sea sediments, and make up approximately 15% of the ocean floor. Oozes are defined as sediments which contain at least 30% skeletal remains of pelagic microorganisms. Siliceous oozes are largely composed of the silica based skeletons of microscopic marine organisms such as diatoms and radiolarians. Other components of siliceous oozes near continental margins may include terrestrially derived silica particles and sponge spicules. Siliceous oozes are composed of skeletons made from opal silica SiO2·nH2O, as opposed to calcareous oozes, which are made from skeletons of calcium carbonate (CaCO3·nH2O) organisms (i.e. coccolithophores). Silica (Si) is a bioessential element and is efficiently recycled in the marine environment through the silica cycle. Distance from land masses, water depth and ocean fertility are all factors that affect the opal silica content in seawater and the presence of siliceous oozes.

Suberites is a genus of sea sponges in the family Suberitidae. Sponges, known scientifically as Porifera, are the oldest metazoans and are used to elucidate the basics of multicellular evolution. These living fossils are ideal for studying the principal features of metazoans, such as extracellular matrix interactions, signal-receptor systems, nervous or sensory systems, and primitive immune systems. Thus, sponges are useful tools with which to study early animal evolution. They appeared approximately 580 million years ago, in the Ediacaran.
Thalassiosira weissflogii is a species of centric diatoms, a unicellular microalga. It is found in marine environments and also in inland waters in many parts of the world. It is actively studied because it may use C4-plant style strategies to increase its photosynthetic efficiency.

Thalassiosirales is an order of centric diatoms. The order currently contains 471 species. Species in the order Thalassiosirales are common in brackish, nearshore, and open-ocean habitats, with approximately the same number of freshwater and marine species.
E. Virginia Armbrust is a biological oceanographer, professor, and current director of the University of Washington School of Oceanography. She is an elected member of the Washington State Academy of Science, an elected fellow of the American Association for the Advancement of Science, and an elected fellow of the American Academy of Microbiology.

Thalassiosiraceae is a family of diatoms in the order Thalassiosirales. The family of Thalassiosiraceae have the unique quality of having a flat valve face. These diatoms are common in brackish, nearshore, and open-ocean habitats, with approximately the same number of freshwater and marine species. Thalassiosiraceae are a centric diatom full of fultoportula. These can often be mistaken for Areola. These belong to many diatom families and can be found in different forms such as the different Areolae that can be found on Navicula or Gomphoneis known as lineolate and punctate. Unlike naviculaceae who are symmetrical in shape some Thalassiosiraceae take on being tangentially undulate.
Thalassiosira is a genus of centric diatoms, comprising over 100 marine and freshwater species. It is a diverse group of photosynthetic eukaryotes that make up a vital part of marine and freshwater ecosystems, in which they are key primary producers and essential for carbon cycling
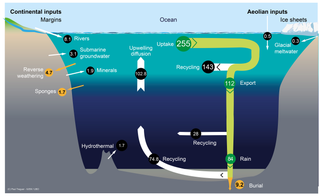
The silica cycle is the biogeochemical cycle in which biogenic silica is transported between the Earth's systems. Silicon is considered a bioessential element and is one of the most abundant elements on Earth. The silica cycle has significant overlap with the carbon cycle and plays an important role in the sequestration of carbon through continental weathering, biogenic export and burial as oozes on geologic timescales.

Marine protists are defined by their habitat as protists that live in marine environments, that is, in the saltwater of seas or oceans or the brackish water of coastal estuaries. Life originated as marine single-celled prokaryotes and later evolved into more complex eukaryotes. Eukaryotes are the more developed life forms known as plants, animals, fungi and protists. Protists are the eukaryotes that cannot be classified as plants, fungi or animals. They are mostly single-celled and microscopic. The term protist came into use historically as a term of convenience for eukaryotes that cannot be strictly classified as plants, animals or fungi. They are not a part of modern cladistics because they are paraphyletic.

Many protists have protective shells or tests, usually made from silica (glass) or calcium carbonate (chalk). Protists are a diverse group of eukaryote organisms that are not plants, animals, or fungi. They are typically microscopic unicellular organisms that live in water or moist environments.
Sonya Dyhrman is an earth and environmental sciences professor who studies the physiology of phytoplankton and their role within marine ecosystems. She is also a fellow of the American Academy of Microbiology.

In geology, silicification is a petrification process in which silica-rich fluids seep into the voids of Earth materials, e.g., rocks, wood, bones, shells, and replace the original materials with silica (SiO2). Silica is a naturally existing and abundant compound found in organic and inorganic materials, including Earth's crust and mantle. There are a variety of silicification mechanisms. In silicification of wood, silica permeates into and occupies cracks and voids in wood such as vessels and cell walls. The original organic matter is retained throughout the process and will gradually decay through time. In the silicification of carbonates, silica replaces carbonates by the same volume. Replacement is accomplished through the dissolution of original rock minerals and the precipitation of silica. This leads to a removal of original materials out of the system. Depending on the structures and composition of the original rock, silica might replace only specific mineral components of the rock. Silicic acid (H4SiO4) in the silica-enriched fluids forms lenticular, nodular, fibrous, or aggregated quartz, opal, or chalcedony that grows within the rock. Silicification happens when rocks or organic materials are in contact with silica-rich surface water, buried under sediments and susceptible to groundwater flow, or buried under volcanic ashes. Silicification is often associated with hydrothermal processes. Temperature for silicification ranges in various conditions: in burial or surface water conditions, temperature for silicification can be around 25°−50°; whereas temperatures for siliceous fluid inclusions can be up to 150°−190°. Silicification could occur during a syn-depositional or a post-depositional stage, commonly along layers marking changes in sedimentation such as unconformities or bedding planes.

Tatiana Rynearson is an American oceanographer who is a professor at the University of Rhode Island. Her research considers plankton diversity and abundance. Rynearson has been on several research cruises, including trips to the North Sea, Puget Sound, the Gulf of Mexico and the North Atlantic.
Greta Albrecht Fryxell was a marine scientist known for her work on the biology and taxonomy of diatoms. In 1996, she was elected a fellow of the American Association for the Advancement of Science.
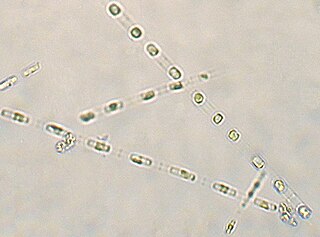
Skeletonema costatum is a cosmopolitan centric diatom that belongs to the genus Skeletonema. It was first described by R. K. Greville, who originally named it Melosira costata, in 1866. It was later renamed by Cleve in 1873 and was more narrowly defined by Zingone et al. and Sarno et al. Skeletonemacostatum is the most well known species of the genus Skeletonema and is often one of the dominant species responsible for red tide events.
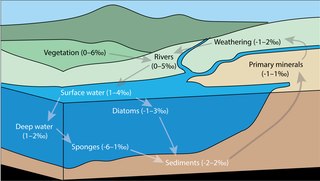
Silicon isotope biogeochemistry is the study of environmental processes using the relative abundance of Si isotopes. As the relative abundance of Si stable isotopes varies among different natural materials, the differences in abundance can be used to trace the source of Si, and to study biological, geological, and chemical processes. The study of stable isotope biogeochemistry of Si aims to quantify the different Si fluxes in the global biogeochemical silicon cycle, to understand the role of biogenic silica within the global Si cycle, and to investigate the applications and limitations of the sedimentary Si record as an environmental and palaeoceanographic proxy.


