A diatom is any member of a large group comprising several genera of algae, specifically microalgae, found in the oceans, waterways and soils of the world. Living diatoms make up a significant portion of the Earth's biomass: they generate about 20 to 50 percent of the oxygen produced on the planet each year, take in over 6.7 billion tonnes of silicon each year from the waters in which they live, and constitute nearly half of the organic material found in the oceans. The shells of dead diatoms can reach as much as a half-mile deep on the ocean floor, and the entire Amazon basin is fertilized annually by 27 million tons of diatom shell dust transported by transatlantic winds from the African Sahara, much of it from the Bodélé Depression, which was once made up of a system of fresh-water lakes.

Phaeodactylum tricornutum is a diatom. It is the only species in the genus Phaeodactylum. Unlike other diatoms, P. tricornutum can exist in different morphotypes and changes in cell shape can be stimulated by environmental conditions. This feature can be used to explore the molecular basis of cell shape control and morphogenesis. Unlike most diatoms, P. tricornutum can grow in the absence of silicon and can survive without making silicified frustules. This provides opportunities for experimental exploration of silicon-based nanofabrication in diatoms.

Asterionella is a genus of pennate freshwater diatoms. They are frequently found in star-shaped colonies of individuals.

Photosynthetic picoplankton or picophytoplankton is the fraction of the photosynthetic phytoplankton of cell sizes between 0.2 and 2 µm. It is especially important in the central oligotrophic regions of the world oceans that have very low concentration of nutrients.
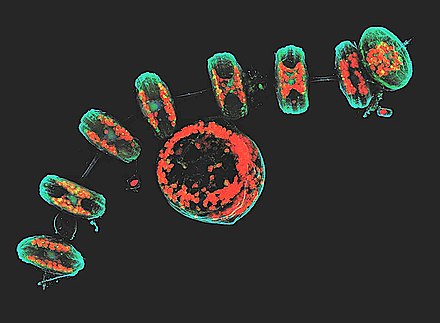
Auxospores are specialised cells in diatoms that are produced at key stages in their cell cycle or life history. Auxospores typically play a role in growth processes, sexual reproduction or dormancy.

Thalassiosira pseudonana is a species of marine centric diatoms. It was chosen as the first eukaryotic marine phytoplankton for whole genome sequencing. T. pseudonana was selected for this study because it is a model for diatom physiology studies, belongs to a genus widely distributed throughout the world's oceans, and has a relatively small genome at 34 mega base pairs. Scientists are researching on diatom light absorption, using the marine diatom of Thalassiosira. The diatom requires a high enough concentration of CO2 in order to utilize C4 metabolism (Clement et al. 2015).

Chaetoceros is a genus of diatoms in the family Chaetocerotaceae, first described by the German naturalist C. G. Ehrenberg in 1844. Species of this genus are mostly found in marine habitats, but a few species exist in freshwater. It is arguably the common and most diverse genus of marine planktonic diatoms, with over 200 accepted species. It is the type genus of its family.

Ochrophytes, also known as heterokontophytes or stramenochromes, are a group of algae. They are the photosynthetic stramenopiles, a group of eukaryotes, organisms with a cell nucleus, characterized by the presence of two unequal flagella, one of which has tripartite hairs called mastigonemes. In particular, they are characterized by photosynthetic organelles or plastids enclosed by four membranes, with membrane-bound compartments called thylakoids organized in piles of three, chlorophyll a and c as their photosynthetic pigments, and additional pigments such as β-carotene and xanthophylls. Ochrophytes are one of the most diverse lineages of eukaryotes, containing ecologically important algae such as brown algae and diatoms. They are classified either as phylum Ochrophyta or Heterokontophyta, or as subphylum Ochrophytina within phylum Gyrista. Their plastids are of red algal origin.
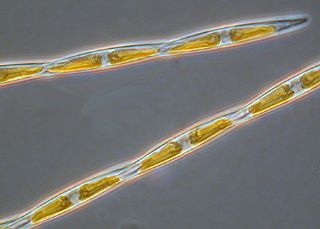
Pseudo-nitzschia is a marine planktonic diatom genus that accounts for 4.4% of pennate diatoms found worldwide. Some species are capable of producing the neurotoxin domoic acid (DA), which is responsible for the neurological disorder in humans known as amnesic shellfish poisoning (ASP). Currently, 58 species are known, 28 of which have been shown to produce DA. It was originally hypothesized that only dinoflagellates could produce harmful algal toxins, but a deadly bloom of Pseudo-nitzschia occurred in 1987 in the bays of Prince Edward Island, Canada, and led to an outbreak of ASP. Over 100 people were affected by this outbreak after consuming contaminated mussels; three people died. Since this event, no additional deaths have been attributed to ASP, though the prevalence of toxic diatoms and DA has increased worldwide. This anomaly is likely due to increased awareness of harmful algal blooms (HABs) and their implications for human and ecosystem health.

Marine microorganisms are defined by their habitat as microorganisms living in a marine environment, that is, in the saltwater of a sea or ocean or the brackish water of a coastal estuary. A microorganism is any microscopic living organism or virus, which is invisibly small to the unaided human eye without magnification. Microorganisms are very diverse. They can be single-celled or multicellular and include bacteria, archaea, viruses, and most protozoa, as well as some fungi, algae, and animals, such as rotifers and copepods. Many macroscopic animals and plants have microscopic juvenile stages. Some microbiologists also classify viruses as microorganisms, but others consider these as non-living.
Thalassiosira weissflogii is a species of centric diatoms, a unicellular microalga. It is found in marine environments and also in inland waters in many parts of the world. It is actively studied because it may use C4-plant style strategies to increase its photosynthetic efficiency.

Ditylum brightwellii is a species of cosmopolitan marine centric diatoms. It is a unicellular photosynthetic autotroph that has the ability to divide rapidly and contribute to spring phytoplankton blooms.

Thalassiosirales is an order of centric diatoms. The order currently contains 471 species. Species in the order Thalassiosirales are common in brackish, nearshore, and open-ocean habitats, with approximately the same number of freshwater and marine species.

Thalassiosiraceae is a family of diatoms in the order Thalassiosirales. The family of Thalassiosiraceae have the unique quality of having a flat valve face. These diatoms are common in brackish, nearshore, and open-ocean habitats, with approximately the same number of freshwater and marine species. Thalassiosiraceae are a centric diatom full of fultoportula. These can often be mistaken for Areola. These belong to many diatom families and can be found in different forms such as the different Areolae that can be found on Navicula or Gomphoneis known as lineolate and punctate. Unlike naviculaceae who are symmetrical in shape some Thalassiosiraceae take on being tangentially undulate.
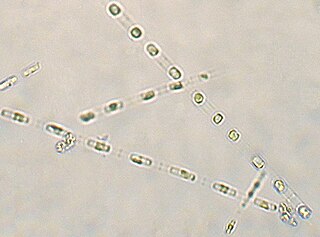
Skeletonema is a genus of diatoms in the family Skeletonemataceae. It is the type genus of its family. The genus Skeletonema was established by R. K. Greville in 1865 for a single species, S. barbadense, found in the Barbados deposit [Jung 2009]. These diatoms are photosynthetic organisms, meaning they obtain carbon dioxide from their surrounding environment and produce oxygen along with other byproducts. Reproduce sexually and asexually [Guiry 2011]. Skeletonema belong to the morphological category referred to as centric diatoms. These are classified by having valves with radial symmetry and the cells lack significant motility [Horner 2002]. Skeletonema are cylindrical shaped with a silica frustule. Cells are joined by long marginal processes to form a filament [Horner 2002]. Their length ranges from 2-61 micrometers, with a diameter ranging from 2-21 micrometers [Hasle 1997]. They are found typically in the neritic zone of the ocean and are highly populous in coastal systems [Jung 2009]. The genus is considered cosmopolitan, showing a wide range of tolerance for salinity and temperature [Hasle 1973]. For example, they have been found in various aquatic environments such as brackish or freshwater. Skeletonema are found worldwide excluding Antarctic waters [Hevia-Orube 2016]. Some harmful effects these diatoms may have on an ecosystem are attributed to large blooming events which may cause hypoxic events in coastal systems. Additionally, they are known to cause water discoloration [Kraberg 2010].
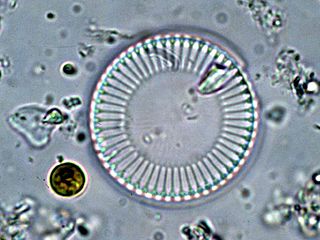
Cyclotella is a genus of diatoms often found in oligotrophic environments, both marine and fresh water. It is in the family Stephanodiscaceae and the order Thalassiosirales. The genus was first discovered in the mid-1800s and since then has become an umbrella genus for nearly 100 different species, the most well-studied and the best known being Cyclotella meneghiniana. Despite being among the most dominant genera in low-productivity environments, it is relatively understudied.

Marine protists are defined by their habitat as protists that live in marine environments, that is, in the saltwater of seas or oceans or the brackish water of coastal estuaries. Life originated as marine single-celled prokaryotes and later evolved into more complex eukaryotes. Eukaryotes are the more developed life forms known as plants, animals, fungi and protists. Protists are the eukaryotes that cannot be classified as plants, fungi or animals. They are mostly single-celled and microscopic. The term protist came into use historically as a term of convenience for eukaryotes that cannot be strictly classified as plants, animals or fungi. They are not a part of modern cladistics because they are paraphyletic.

Many protists have protective shells or tests, usually made from silica (glass) or calcium carbonate (chalk). Protists are a diverse group of eukaryote organisms that are not plants, animals, or fungi. They are typically microscopic unicellular organisms that live in water or moist environments.

Tatiana Rynearson is an American oceanographer who is a professor at the University of Rhode Island. Her research considers plankton diversity and abundance. Rynearson has been on several research cruises, including trips to the North Sea, Puget Sound, the Gulf of Mexico and the North Atlantic.
Algal viruses are the viruses infecting algae, which are photosynthetic single-celled eukaryotes. As of 2020, there were 61 viruses known to infect algae. Algae are integral components of aquatic food webs and drive nutrient cycling, so the viruses infecting algal populations also impacts the organisms and nutrient cycling systems that depend on them. Thus, these viruses can have significant, worldwide economic and ecological effects. Their genomes varied between 4.4 to 560 kilobase pairs (kbp) long and used double-stranded Deoxyribonucleic Acid (dsDNA), double-stranded Ribonucleic Acid (dsRNA), single-stranded Deoxyribonucleic Acid (ssDNA), and single-stranded Ribonucleic Acid (ssRNA). The viruses ranged between 20 and 210 nm in diameter. Since the discovery of the first algae-infecting virus in 1979, several different techniques have been used to find new viruses infecting algae and it seems that there are many algae-infecting viruses left to be discovered