| Hyphomycetes | |
|---|---|
| Scientific classification | |
| Kingdom: | |
| Orders | |
Hyphomycetales (obsolete) Contents | |
Hyphomycetes are a form classification of fungi, part of what has often been referred to as fungi imperfecti, Deuteromycota, or anamorphic fungi. Hyphomycetes lack closed fruit bodies, and are often referred to as moulds (or molds). Most hyphomycetes are now assigned to the Ascomycota, on the basis of genetic connections made by life-cycle studies or by phylogenetic analysis of DNA sequences; many remain unassigned phylogenetically.
Although no longer considered a phylogenetically defined taxon, the prevalence of hyphomycete forms in nature, the built environment, and laboratories means that identification of members this group remains of practical importance.
Because asexual forms of fungi usually occur separately from their sexual forms, when microscopic fungi began to be studied in the early 19th century, it was often unknown when two morphologically different forms were actually part of one species. The tendency for some organisms to apparently only have asexual forms, or for their sexual forms to be discovered long after the asexual forms, meant that an independent taxonomy was developed for asexual fungi. Near the beginning of the 20th century, when it became clearer that many asexual and sexual forms were related, the concept of 'form taxa' was developed. The independent taxonomy of asexual forms was regarded as artificial, not representative of evolutionary relationships, and intended to be practical for identification purposes. The taxonomy of the sexual states was considered the true classification. The result was that many fungal species ended up with two accepted Latin binomials, one for the asexual form (or anamorph) and the other for the sexual form (teleomorph). This dual nomenclature was only abandoned in January 2012, [1] and the transition to a single name system, with one name representing all morphs of a fungus, is still incomplete. [2]
Traditional identification of hyphomycetes was primarily based on microscopic morphology including: conidial morphology, especially septation, shape, size, colour and cell wall texture, the arrangement of conidia as they are borne on the conidiogenous cells (e.g. if they are solitary, in chains, or produced in slime), the type of conidiogenous cell (e.g. non-specialized or hypha-like, phialide, annellide, or sympodial), and other additional features such as the presence of sporodochia or synnemata. [3] [4] [5]
For species growing in culture, or in environmental DNA studies, most identifications of Hyphomycetes are now done with DNA barcoding. This is not always possible, however, for archival specimens or samples such as microscopic slides from air samples.
Aquatic or Ingoldian hyphomycetes are common on submerged decaying leaves and other organic matter, especially in clean running water with good aeration. Colonised leaves fall from the tree into the river. Their branched, septate mycelium penetrates through the leaf surface and spreads through leaf tissue. Conidiophores project into the water and bear conidia, which are often sigmoid, branched or tetraradiate structures. Aquatic hyphomycetes play an important role in the breakdown of organic matter in rivers, because their extracellular enzymes break down leaf tissue, which in turn is made more palatable to invertebrates. Leaves with fungi (conditioned) are a more nutritious source of food than unconditioned leaves. [6]
Coprophilous or dung-loving hyphomycetes are part of the succession of fungi occurring on many kinds of herbivore faeces, playing an important role in breaking down cellulose. [7] Several species are found only on dung, such as Angulimaya sundara , Onychophora coprophila , Pulchromyces fimicola , Sphondylocephalum verticillatum and Stilbella fimetaria .
Entomogenous, entomopathogenic or insect-pathogenic hyphomycetes infect and kill insects (and spiders) and are especially diverse in tropical and subtropical regions, especially in Asia. [8] Most are asexual states of or phylogenetically related to the Ascomycete families, Cordycipitaceae and Ophiocordycipitaceae. Insect hosts are infected by asexual spores, which germinate and grow to fill the host body with mycelium or hyphal bodies, then produce sporulating structures on the insect carcass. They are often found on dead insects under bark or in soil, but some affect insect behaviour ("zombie fungus"), causing infected hosts to climb towards the light, ensuring that air-borne infective spores will be released higher up in the canopy of the forest or meadow. [9] Well-known entomogenous hyphomycetes are classified in Beauveria , Metarhizium and Tolypocladium ; famous anamorphic generic names such as Akanthomyces , Gibellula , Hirsutella , Hymenostilbe and Isaria are now subsumed in genera formerly considered sexual, such as Cordyceps , Ophiocordyceps and Torubiella under fungal single-name nomenclature. [10] Species of Beauveria and Metarhizium show some promise as biological control agents against pest insects. [11] Tolypocladium inflatum was the original source of cyclosporine A, used as a drug to prevent rejection of organ transplants. [12]
Many food-borne fungi are hyphomycetes. Species of Penicillium and Aspergillus are particularly common agents of food spoilage and also produce important mycotoxins that affect human health. [13] Some species, such as Penicillium digitatum on citrus fruits, and Penicillium expansum on apples, are common on specific foods, while others are less specialized and grow on many different kinds of food.
Nematophagous or nematode-trapping hyphomycetes either live their life-cycles in the bodies of dead nematodes or trap and kill nematodes in order to supplement their nitrogen requirements. [14] Species of the hyphomycete genus Arthrobotrys , phylogenetically related to or being the asexual forms of Orbilia , produce constricting loops that quickly shut to grab nematodes, or non-constricting loops or hyphal networks that entangle nematodes, or sticky knobs that adhere to nematodes as they swim by. Attempts to exploit these fungi as biological control agents against agriculturally harmful nematodes have generally been unsuccessful. [15]

Ascomycota is a phylum of the kingdom Fungi that, together with the Basidiomycota, forms the subkingdom Dikarya. Its members are commonly known as the sac fungi or ascomycetes. It is the largest phylum of Fungi, with over 64,000 species. The defining feature of this fungal group is the "ascus", a microscopic sexual structure in which nonmotile spores, called ascospores, are formed. However, some species of the Ascomycota are asexual, meaning that they do not have a sexual cycle and thus do not form asci or ascospores. Familiar examples of sac fungi include morels, truffles, brewers' and bakers' yeast, dead man's fingers, and cup fungi. The fungal symbionts in the majority of lichens such as Cladonia belong to the Ascomycota.

Penicillium is a genus of ascomycetous fungi that is part of the mycobiome of many species and is of major importance in the natural environment, in food spoilage, and in food and drug production.

Beauveria bassiana is a fungus that grows naturally in soils throughout the world and acts as a parasite on various arthropod species, causing white muscardine disease; it thus belongs to the group of entomopathogenic fungi. It is used as a biological insecticide to control a number of pests, including termites, thrips, whiteflies, aphids and various beetles. Its use in the control of bedbugs and malaria-transmitting mosquitos is under investigation.
An entomopathogenic fungus is a fungus that can kill or seriously disable insects.

Metarhizium robertsii – formerly known as M. anisopliae, and even earlier as Entomophthora anisopliae (basionym) – is a fungus that grows naturally in soils throughout the world and causes disease in various insects by acting as a parasitoid. Ilya I. Mechnikov named it after the insect species from which it was originally isolated – the beetle Anisoplia austriaca. It is a mitosporic fungus with asexual reproduction, which was formerly classified in the form class Hyphomycetes of the phylum Deuteromycota.
Tolypocladium inflatum is an ascomycete fungus originally isolated from a Norwegian soil sample that, under certain conditions, produces the immunosuppressant drug ciclosporin. In its sexual stage (teleomorph) it is a parasite on scarab beetles. It forms a small, compound ascocarp that arises from the cadaver of its host beetle. In its asexual stage (anamorph) it is a white mold that grows on soil. It is much more commonly found in its asexual stage and this is the stage that was originally given the name Tolypocladium inflatum.

Epichloë is a genus of ascomycete fungi forming an endophytic symbiosis with grasses. Grass choke disease is a symptom in grasses induced by some Epichloë species, which form spore-bearing mats (stromata) on tillers and suppress the development of their host plant's inflorescence. For most of their life cycle however, Epichloë grow in the intercellular space of stems, leaves, inflorescences, and seeds of the grass plant without incurring symptoms of disease. In fact, they provide several benefits to their host, including the production of different herbivore-deterring alkaloids, increased stress resistance, and growth promotion.

Beauveria is a genus of asexually-reproducing fungi allied with the ascomycete family Cordycipitaceae. Its several species are typically insect pathogens. The sexual states (teleomorphs) of Beauveria species, where known, are species of Cordyceps.

Ophiocordyceps unilateralis, commonly known as zombie-ant fungus, is an insect-pathogenic fungus, discovered by the British naturalist Alfred Russel Wallace in 1859, and currently found predominantly in tropical forest ecosystems. O. unilateralis infects ants of the tribe Camponotini, with the full pathogenesis being characterized by alteration of the behavioral patterns of the infected ant. Infected hosts leave their canopy nests and foraging trails for the forest floor, an area with a temperature and humidity suitable for fungal growth; they then use their mandibles to attach themselves to a major vein on the underside of a leaf, where the host remains after its eventual death. The process, leading up to mortality, takes 4–10 days, and includes a reproductive stage where fruiting bodies grow from the ant's head, rupturing to release the fungus's spores. O. unilateralis is, in turn, also susceptible to fungal infection itself, an occurrence that can limit its impact on ant populations, which has otherwise been known to devastate ant colonies.

A fungus is any member of the group of eukaryotic organisms that includes microorganisms such as yeasts and molds, as well as the more familiar mushrooms. These organisms are classified as one of the traditional eukaryotic kingdoms, along with Animalia, Plantae and either Protista or Protozoa and Chromista.

The Orbiliaceae are a family of saprobic sac fungi in the order Orbiliales. The family, first described by John Axel Nannfeldt in 1932, contains 288 species in 12 genera. Members of this family have a widespread distribution, but are more prevalent in temperate regions. Some species in the Orbiliaceae are carnivorous fungi, and have evolved a number of specialized mechanisms to trap nematodes.

Metarhizium flavoviride is a Sordariomycete in the order Hypocreales and family Clavicipitaceae. The genus Metarhizium currently consists of 45 described species and are a group of fungal isolates that are known to be virulent against Hemiptera and some Coleoptera. M. flavoviride is described as its own species, but there also exists a variety of M. flavoviride, which is M. flavoviride var. flavoviride. Previously described varieties of M. flavoviride have been documented, however recent random amplified polymorphic DNA (RAPD) markers have assigned these varieties as new species. The reassigned species are as follows: M. flavoviride Type E is now M. brasiliense; M. flavoviride var. minus is now M. minus; M. flavoviride var. novozealandicum is now M. novozealandicum; and M. flavoviride var. pemphigi is now M. pemphigi.
Podonectria is a genus in the monotypic Podonectriaceae family of fungi. They are parasitic fungus on scale insects, other fungi, or on substrates that had previously colonized by other fungi.
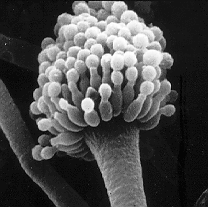
The fungi imperfecti or imperfect fungi are fungi which do not fit into the commonly established taxonomic classifications of fungi that are based on biological species concepts or morphological characteristics of sexual structures because their sexual form of reproduction has never been observed. They are known as imperfect fungi because only their asexual and vegetative phases are known. They have asexual form of reproduction, meaning that these fungi produce their spores asexually, in the process called sporogenesis.

Purpureocillium is a fungal genus in the Ophiocordycipitaceae family. The genus now contains at least 5 species with the type species Purpureocillium lilacinum, a common saprobic, filamentous fungus. It has been isolated from a wide range of habitats, including cultivated and uncultivated soils, forests, grassland, deserts, estuarine sediments and sewage sludge, and insects. It has also been found in nematode eggs, and occasionally from females of root-knot and cyst nematodes. In addition, it has frequently been detected in the rhizosphere of many crops. The species can grow at a wide range of temperatures – from 8 to 38 °C for a few isolates, with optimal growth in the range 26 to 30 °C. It also has a wide pH tolerance and can grow on a variety of substrates. P. lilacinum has shown promising results for use as a biocontrol agent to control the growth of destructive root-knot nematodes.
Purpureocillium lilacinum is a species of filamentous fungus in the family Ophiocordycipitaceae. It has been isolated from a wide range of habitats, including cultivated and uncultivated soils, forests, grassland, deserts, estuarine sediments and sewage sludge, and insects. It has also been found in nematode eggs, and occasionally from females of root-knot and cyst nematodes. In addition, it has frequently been detected in the rhizosphere of many crops. The species can grow at a wide range of temperatures – from 8 to 38 °C for a few isolates, with optimal growth in the range 26 to 30 °C. It also has a wide pH tolerance and can grow on a variety of substrates. P. lilacinum has shown promising results for use as a biocontrol agent to control the growth of destructive root-knot nematodes.

Tolypocladium is a genus of fungi within the family Ophiocordycipitaceae. It includes species that are parasites of other fungi, insect pathogens, rotifer pathogens and soil inhabiting species with uncertain ecological roles. Tolypocladium was originally circumscribed as a genus containing anamorphic fungi. It was later determined that some Cordyceps-like teleomorphic fungi were the teleomorphs of Tolypocladium species. These species were considered to belong in the genus Cordyceps until molecular phylogenetics studies found these species to be more closely related to Ophiocordyceps and were considered to belong in that genus before they were transferred to the new genus Elaphocordyceps by Sung and colleagues in 2007. However, under the ICN's 2011 "one fungus, one name" principle, fungi can not have different names for their anamorphic and teleomorphic stages if they are found to be the same taxon. Quandt and colleagues formally synonymized Tolypocladium and Elaphocordyceps in 2014. Quandt and colleagues also synonymized the anamorphic genus Chaunopycnis with Tolypocladium. The immunosuppressant drug ciclosporin was originally isolated from Tolypocladium inflatum, and has since been found in other species of Tolypocladium, some of which were formerly placed in Chaunopycnis.
Muscardine is a disease of insects. It is caused by many species of entomopathogenic fungus. Many muscardines are known for affecting silkworms. Muscardine may also be called calcino.
Donald W. Roberts was an American insect pathologist and one of the originators of that field. He was especially known for research into biological pest control of Lepidoptera by Metarhizium but also Beauveria bassiana. He was a Research Professor Emeritus in the Biology Department of Utah State University.

Cordyceps locustiphila is the basionym and teleomorph of the fungi Beauveria locustiphila, a species of fungus in the family Cordycipitaceae. and is a species within the genus Cordyceps. It was originally described in by Henn in 1904. C. locustiphila is an entomopathogen and obligate parasite of the grasshopper species within the genus Colpolopha or Tropidacris, and as such is endemic to South America. The scientific name is derived from its close relationship with its host, being named after locusts. The fungi was renamed to Beauveria locustiphila in 2017 following research into the family Cordycipitaceae. Following the loss of the species type specimen, new studies were conducted that now recommend that the fungi be divided into 3 species. C. locustiphila, C. diapheromeriphila, and C. acridophila.
{{cite book}}: |journal= ignored (help)