Ardipithecus is a genus of an extinct hominine that lived during the Late Miocene and Early Pliocene epochs in the Afar Depression, Ethiopia. Originally described as one of the earliest ancestors of humans after they diverged from the chimpanzees, the relation of this genus to human ancestors and whether it is a hominin is now a matter of debate. Two fossil species are described in the literature: A. ramidus, which lived about 4.4 million years ago during the early Pliocene, and A. kadabba, dated to approximately 5.6 million years ago. Initial behavioral analysis indicated that Ardipithecus could be very similar to chimpanzees, however more recent analysis based on canine size and lack of canine sexual dimorphism indicates that Ardipithecus was characterised by reduced aggression, and that they more closely resemble bonobos.

Bipedalism is a form of terrestrial locomotion where a tetrapod moves by means of its two rear limbs or legs. An animal or machine that usually moves in a bipedal manner is known as a biped, meaning 'two feet'. Types of bipedal movement include walking or running and hopping.

Homininae, also called "African hominids" or "African apes", is a subfamily of Hominidae. It includes two tribes, with their extant as well as extinct species: 1) the tribe Hominini ―and 2) the tribe Gorillini (gorillas). Alternatively, the genus Pan is sometimes considered to belong to its own third tribe, Panini. Homininae comprises all hominids that arose after orangutans split from the line of great apes. The Homininae cladogram has three main branches, which lead to gorillas and to humans and chimpanzees. There are two living species of Panina and two living species of gorillas, but only one extant human species. Traces of extinct Homo species, including Homo floresiensis have been found with dates as recent as 40,000 years ago. Organisms in this subfamily are described as hominine or hominines.

Australopithecus is a genus of early hominins that existed in Africa during the Pliocene and Early Pleistocene. The genera Homo, Paranthropus, and Kenyanthropus evolved from some Australopithecus species. Australopithecus is a member of the subtribe Australopithecina, which sometimes also includes Ardipithecus, though the term "australopithecine" is sometimes used to refer only to members of Australopithecus. Species include A. garhi, A. africanus, A. sediba, A. afarensis, A. anamensis, A. bahrelghazali and A. deyiremeda. Debate exists as to whether some Australopithecus species should be reclassified into new genera, or if Paranthropus and Kenyanthropus are synonymous with Australopithecus, in part because of the taxonomic inconsistency.

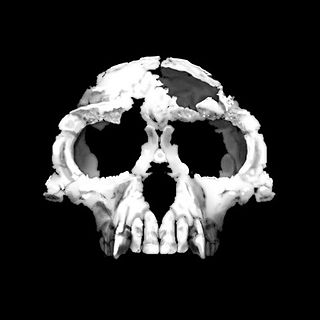
Sahelanthropus tchadensis is an extinct species of hominid dated to about 7 million years ago, during the Miocene epoch. The species, and its genus Sahelanthropus, was announced in 2002, based mainly on a partial cranium, nicknamed Toumaï, discovered in northern Chad.

Australopithecus afarensis is an extinct species of australopithecine which lived from about 3.9–2.9 million years ago (mya) in the Pliocene of East Africa. The first fossils were discovered in the 1930s, but major fossil finds would not take place until the 1970s. From 1972 to 1977, the International Afar Research Expedition—led by anthropologists Maurice Taieb, Donald Johanson and Yves Coppens—unearthed several hundreds of hominin specimens in Hadar, Ethiopia, the most significant being the exceedingly well-preserved skeleton AL 288-1 ("Lucy") and the site AL 333. Beginning in 1974, Mary Leakey led an expedition into Laetoli, Tanzania, and notably recovered fossil trackways. In 1978, the species was first described, but this was followed by arguments for splitting the wealth of specimens into different species given the wide range of variation which had been attributed to sexual dimorphism. A. afarensis probably descended from A. anamensis and is hypothesised to have given rise to Homo, though the latter is debated.

Oreopithecus is an extinct genus of hominoid primate from the Miocene epoch whose fossils have been found in today's Tuscany and Sardinia in Italy. It existed nine to seven million years ago in the Tusco-Sardinian area when this region was an isolated island in a chain of islands stretching from central Europe to northern Africa in what was becoming the Mediterranean Sea.

Brachiation, or arm swinging, is a form of arboreal locomotion in which primates swing from tree limb to tree limb using only their arms. During brachiation, the body is alternately supported under each forelimb. This form of locomotion is the primary means of locomotion for the small gibbons and siamangs of southeast Asia. Gibbons in particular use brachiation for as much as 80% of their locomotor activities. Some New World monkeys, such as spider monkeys and muriquis, were initially classified as semibrachiators and move through the trees with a combination of leaping and brachiation. Some New World species also practice suspensory behaviors by using their prehensile tail, which acts as a fifth grasping hand. Evidence has shown that the extinct ape Proconsul from the Miocene of East Africa developed an early form of suspensory behaviour, and was therefore referred to as a probrachiator.

Australopithecus africanus is an extinct species of australopithecine which lived between about 3.3 and 2.1 million years ago in the Late Pliocene to Early Pleistocene of South Africa. The species has been recovered from Taung, Sterkfontein, Makapansgat, and Gladysvale. The first specimen, the Taung child, was described by anatomist Raymond Dart in 1924, and was the first early hominin found. However, its closer relations to humans than to other apes would not become widely accepted until the middle of the century because most had believed humans evolved outside of Africa. It is unclear how A. africanus relates to other hominins, being variously placed as ancestral to Homo and Paranthropus, to just Paranthropus, or to just P. robustus. The specimen "Little Foot" is the most completely preserved early hominin, with 90% of the skeleton intact, and the oldest South African australopith. However, it is controversially suggested that it and similar specimens be split off into "A. prometheus".
Orthograde is a term derived from Greek ὀρθός, orthos + Latin gradi that describes a manner of walking which is upright, with the independent motion of limbs. Both New and Old World monkeys are primarily arboreal, and they have a tendency to walk with their limbs swinging in parallel to one another. This differs from the manner of walking demonstrated by the apes.
A facultative biped is an animal that is capable of walking or running on two legs (bipedal), as a response to exceptional circumstances (facultative), while normally walking or running on four limbs or more. In contrast, obligate bipedalism is where walking or running on two legs is the primary method of locomotion. Facultative bipedalism has been observed in several families of lizards and multiple species of primates, including sifakas, capuchin monkeys, baboons, gibbons, gorillas, bonobos and chimpanzees. Several dinosaur and other prehistoric archosaur species are facultative bipeds, most notably ornithopods and marginocephalians, with some recorded examples within sauropodomorpha. Different facultatively bipedal species employ different types of bipedalism corresponding to the varying reasons they have for engaging in facultative bipedalism. In primates, bipedalism is often associated with food gathering and transport. In lizards, it has been debated whether bipedal locomotion is an advantage for speed and energy conservation or whether it is governed solely by the mechanics of the acceleration and lizard's center of mass. Facultative bipedalism is often divided into high-speed (lizards) and low-speed (gibbons), but some species cannot be easily categorized into one of these two. Facultative bipedalism has also been observed in cockroaches and some desert rodents.
AL 288-1, commonly known as Lucy or Dinkʼinesh, is a collection of several hundred pieces of fossilized bone comprising 40 percent of the skeleton of a female of the hominin species Australopithecus afarensis. It was discovered in 1974 in Ethiopia, at Hadar, a site in the Awash Valley of the Afar Triangle, by Donald Johanson, a paleoanthropologist of the Cleveland Museum of Natural History.

Ardipithecus ramidus is a species of australopithecine from the Afar region of Early Pliocene Ethiopia 4.4 million years ago (mya). A. ramidus, unlike modern hominids, has adaptations for both walking on two legs (bipedality) and life in the trees (arboreality). However, it would not have been as efficient at bipedality as humans, nor at arboreality as non-human great apes. Its discovery, along with Miocene apes, has reworked academic understanding of the chimpanzee–human last common ancestor from appearing much like modern-day chimpanzees, orangutans and gorillas to being a creature without a modern anatomical cognate.

The evolution of human bipedalism, which began in primates approximately four million years ago, or as early as seven million years ago with Sahelanthropus, or approximately twelve million years ago with Danuvius guggenmosi, has led to morphological alterations to the human skeleton including changes to the arrangement, shape, and size of the bones of the foot, hip, knee, leg, and the vertebral column. These changes allowed for the upright gait to be overall more energy efficient in comparison to quadrupeds. The evolutionary factors that produced these changes have been the subject of several theories that correspond with environmental changes on a global scale.

A hand is a prehensile, multi-fingered appendage located at the end of the forearm or forelimb of primates such as humans, chimpanzees, monkeys, and lemurs. A few other vertebrates such as the koala are often described as having "hands" instead of paws on their front limbs. The raccoon is usually described as having "hands" though opposable thumbs are lacking.
The endurance running hypothesis is a series of conjectures which presume humans evolved anatomical and physiological adaptations to run long distances and, more strongly, that "running is the only known behavior that would account for the different body plans in Homo as opposed to apes or australopithecines".
Ardi (ARA-VP-6/500) is the designation of the fossilized skeletal remains of an Ardipithecus ramidus, thought to be an early human-like female anthropoid 4.4 million years old. It is the most complete early hominid specimen, with most of the skull, teeth, pelvis, hands and feet, more complete than the previously known Australopithecus afarensis specimen called "Lucy". In all, 125 different pieces of fossilized bone were found.
Vertical clinging and leaping (VCL) is a type of arboreal locomotion seen most commonly among the strepsirrhine primates and haplorrhine tarsiers. The animal begins at rest with its torso upright and elbows fixed, with both hands clinging to a vertical support, such as the side of a tree or bamboo stalk. To move from one support to another, it pushes off from one vertical support with its hindlimbs, landing on another vertical support after an extended period of free flight. Vertical clinging and leaping primates have evolved a specialized anatomy to compensate for the physical implications of this form of locomotion. These key morphological specializations have been identified in prosimian fossils from as early as the Eocene.
The savannah hypothesis is a hypothesis that human bipedalism evolved as a direct result of human ancestors' transition from an arboreal lifestyle to one on the savannas. According to the hypothesis, hominins left the woodlands that had previously been their natural habitat millions of years ago and adapted to their new habitat by walking upright.
Danuvius guggenmosi was an extinct species of great ape that lived 11.6 million years ago during the Middle–Late Miocene in southern Germany. It is the sole member of the genus Danuvius. The area at this time was probably a woodland with a seasonal climate. A male specimen was estimated to have weighed about 31 kg (68 lb), and two females 17 and 19 kg. Both genus and species were described in November 2019.
