
Temnospondyli or temnospondyls is a diverse ancient order of small to giant tetrapods—often considered primitive amphibians—that flourished worldwide during the Carboniferous, Permian and Triassic periods, with fossils being found on every continent. A few species continued into the Jurassic and Early Cretaceous periods, but all had gone extinct by the Late Cretaceous. During about 210 million years of evolutionary history, they adapted to a wide range of habitats, including freshwater, terrestrial, and even coastal marine environments. Their life history is well understood, with fossils known from the larval stage, metamorphosis and maturity. Most temnospondyls were semiaquatic, although some were almost fully terrestrial, returning to the water only to breed. These temnospondyls were some of the first vertebrates fully adapted to life on land. Although temnospondyls are amphibians, many had characteristics such as scales and armour-like bony plates that distinguish them from the modern soft-bodied lissamphibians.

Batrachotomus is a genus of prehistoric archosaur. Fossils of this animal have been found in southern Germany and dated from the Ladinian stage of the Middle Triassic period, around 242 to 237 million years ago. Batrachotomus was described by palaeontologist David J. Gower 22 years after its discovery.

Cyclotosaurus is an extinct genus of temnospondyl within the family Mastodonsauridae. It was of great size for an amphibian, had an elongated skull up to 56 cm (22 in).

Uranocentrodon is an extinct genus of temnospondyls in the family Rhinesuchidae. Known from a 50 centimetres (20 in) skull, Uranocentrodon was a large predator with a length up to 3.75 metres (12.3 ft). Originally named Myriodon by van Hoepen in 1911, it was transferred to a new genus on account of the name being preoccupied in 1917. It has been synonymized with Rhinesuchus, but this has not been widely supported. It was also originally considered to be of Triassic age, but more recent analysis has placed its age as just below the Permian-Triassic boundary.
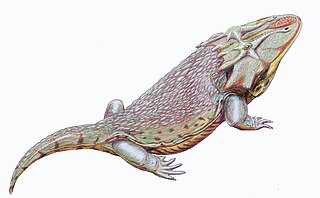
Dasyceps is an extinct genus of zatracheidid temnospondyl from the early Permian of England.

Eocyclotosaurus is an extinct genus of mastodonsauroid temnospondyl from the Middle Triassic (Anisian). The name Eocyclotosaurus means "dawn round-eared lizard". It is characterized as a capitosauroid with a long and slender snout, closed otic fenestra, and small orbits. It measured over one metre and had a 22 cm skull.

Konzhukovia is an amphibian genus that belongs to an extinct family Konzhukoviidae of temnospondyls, the largest clade of basal tetrapods including about 198 genera, 292 species, and more than half of which were alive during the early Mesozoic period. The animal was a predator that lived about 260 million years ago, and could get up to about three meters in length. Specifically, Konzukovia lived during the Permian, between 252 and 270 million years ago according to the type of rock the fossil was found in. There are three species within this genus, K. vetusta, K. tarda, and K. sangabrielensis, the first two originating from Russia while the latest originating from Southern Brazil. The discovery of this specimen in Southern Brazil provided more evidence to support the idea that during this animals existence, there was a “biological corridor” because of the supercontinent Pangea, allowing these species to be found so far apart from each other. Konzhukovia belongs to the family Archegosauridae, a family consisted of large temnospondyls that most likely compare to modern day crocodiles. Since the discovery of the latest species, K. sangabrielensis, Pacheco proposes that there must be the creation of a new family, Konzhokoviidae, a monophyletic group in a sister-group relationship with Stereospondlyi in order to accommodate the three species. Konzhukovia skulls usually exhibit typical rhinesuchid features including an overall parabolic shape, small orbits located more posteriorly, and the pterygoids do not reach the vomer. These animals were long-snouted amphibians that had clear adaptations made for fish catching, as well as exemplifying aquatic features.
Mastodonsauridae is a family of capitosauroid temnospondyls. Fossils belonging to this family have been found in North America, Greenland, Europe, Asia, and Australia. The family Capitosauridae is synonymous with Mastodonsauridae.

The Mastodonsauroidea are an extinct superfamily of temnospondyl amphibians known from the Triassic. Fossils belonging to this superfamily have been found in North America, Greenland, Europe, Asia, and Australia. Ferganobatrachus from the Jurassic of Asia was initially assigned to this superfamily, but is now considered to be congeneric with the brachyopid Gobiops.
Bukobaja is an extinct genus of mastodonsaurid temnospondyl from the middle Triassic of Russia. It contains a single species, Bukobaja enigmatica. Bukobaja mainly occurs in the Bukobay Svita as part of the Ladinian?-age "Mastodonsaurus fauna", a section of Russian Triassic biostratigraphy characterized by "Mastodonsaurus" torvus. It was also present in the underlying Donguz Svita. Bukobaja appears to be a valid genus similar to, yet distinct from, Mastodonsaurus. Despite appearing to possess several unique features, Bukobaja is still known from very few remains. This has led to difficulties in determining its relations more precisely than "Mastodonsauridae incertae sedis". It has also been compared to trematosaurids.

Cryobatrachus is an extinct genus of temnospondyl amphibian from the Early Triassic of Antarctica. The type species is Cryobatrachus kitchingi. It is known from a partial skull and an imprint of the skull roof, both found in the Fremouw Formation of the Transantarctic Mountains at about 85° south latitude and described in 1974. Many small bone fragments have also been identified, although they cannot be attributed with certainty to C. kitchingi. Cryobatrachus has been classified in the family Lydekkerinidae, as it is similar in appearance to the genus Lydekkerina from South Africa. Because only a small number of features distinguish it from other lydekkerinids, Cryobatrachus kitchingi has more recently been considered a nomen dubium, meaning that its distinction from other better-known species may be unwarranted.

Xenotosuchus is an extinct genus of mastodonsaurid temnospondyl within the family Mastodonsauridae known from the Triassic of South Africa. The genus is based on a skull originally described as Parotosuchus, an animal which it resembled in general build and habit.

Plagiosuchus is an extinct genus of plagiosaurid temnospondyl. It is known from several collections from the Middle Triassic of Germany.

Micropholis is an extinct genus of dissorophoid temnospondyl. Fossils have been found from the Lystrosaurus Assemblage Zone of the Karoo Basin in South Africa and are dated to the Induan. Fossils have also been found from the lower Fremouw of Antarctica.Micropholis is the only post-Permian dissorophoid and the only dissorophoid in what is presently the southern hemisphere and what would have been termed Gondwana during the amalgamation of Pangea.
Heptasaurus is an extinct genus of Triassic capitosaurian temnospondyl within the family Mastodonsauridae.

The Erfurt Formation, also known as the Lower Keuper, is a stratigraphic formation of the Keuper group and the Germanic Trias supergroup. It was deposited during the Ladinian stage of the Triassic period. It lies above the Upper Muschelkalk and below the Middle Keuper.
Jaxtasuchus is an extinct genus of armored doswelliid archosauriform reptile known from the Middle Triassic of the Erfurt Formation in Germany. The type species, Jaxtasuchus salomoni, was named in 2013 on the basis of several incomplete skeletons and other isolated remains. Like other doswelliids, members of the genus were heavily armored, with four longitudinal rows of bony plates called osteoderms covering the body. Jaxtasuchus is the first doswelliid known from Europe and is most closely related to Doswellia from the Late Triassic of the eastern United States. However, it was not as specialized as Doswellia, retaining several generalized archosauriform characteristics and having less armor. Jaxtasuchus fossils have been found in aquatic mudstones alongside fossils of temnospondyl amphibians, crustaceans, and mollusks, suggesting that Jaxtasuchus was semiaquatic like modern crocodilians.

Warrenisuchus is an extinct genus of temnospondyl from the Early Triassic of Queensland, Australia. It belongs to a diverse group of Triassic temnospondyls called Capitosauria. The type species Warrenisuchus aliciae was erected in 2009. W. aliciae was originally described as a species of Parotosuchus in 1988, which is known from other species that have been found in Europe, Africa, and Antarctica. In 2000 it was then assigned to a new genus called Rewanobatrachus along with the newly named species R. gunganj, which was declared the type species of the genus. However, R. gunganj was later reclassified as a species of Watsonisuchus, invalidating the name Rewanobatrachus and requiring that R. aliciae be placed in its own genus, which was named Warrenisuchus. However, several studies suggest that Warrenisuchus aliciae may be a species of Watsonisuchus as well. Unlike most capitosaurs, Warrenisuchus is known from many juvenile skulls less than 4 centimetres (1.6 in) in length.
Scapanops is an extinct genus of dissorophid temnospondyl amphibian known from the Early Permian Nocona Formation of north-central Texas, United States. It contains only the type species Scapanops neglecta, which was named by Rainer R. Schoch and Hans-Dieter Sues in 2013. Scapanops differs from other dissorophids in having a very small skull table, which means that its eye sockets are unusually close to the back of the skull. The eye sockets are also very large and spaced far apart. Scapanops was probably small-bodied with a proportionally large head and short trunk and tail. Like other dissorophids, it probably spent most of its life on land.

Manubrantlia was a genus of lapillopsid temnospondyls from the Early Triassic Panchet Formation of India. This genus is only known from a single holotype left jaw, given the designation ISI A 57. Despite the paucity of remains, the jaw is still identifiable as belonging to a relative of Lapillopsis. For example, all three of its coronoid bones possessed teeth, the articular bone is partially visible in lateral (outer) view, and its postsplenial does not contact the posterior meckelian foramen. However, the jaw also possesses certain unique features which justify the erection of a new genus separate from Lapillopsis. For example, the mandible is twice the size of any jaws referred to other lapillopsids. The most notable unique feature is an enlarged "pump-handle" shaped arcadian process at the back of the jaw. This structure is responsible for the generic name of this genus, as "Manubrantlia" translates from Latin to the English expression "pump-handle". The type and only known species of this genus is Manubrantlia khaki. The specific name refers to the greenish-brown mudstones of the Panchet Formation, with a color that had been described as "khaki" by the first British geologists who studied the formation.












![19th century illustration of the complex inner structure of a Labyrinthodon [Mastodonsaurus] tooth Vestiges 11 fig 39 Labyrinthodon tooth.jpg](http://upload.wikimedia.org/wikipedia/commons/thumb/1/1c/Vestiges_11_fig_39_Labyrinthodon_tooth.jpg/171px-Vestiges_11_fig_39_Labyrinthodon_tooth.jpg)

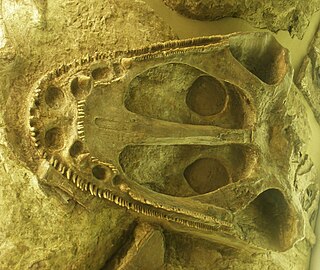
![Underside of the skull of Mastodonsaurus [Heptasaurus] cappelensis in the Naturmuseum Senckenberg in Frankfurt am Main in Germany Heptasaurus kappelensis.jpg](http://upload.wikimedia.org/wikipedia/commons/thumb/6/6a/Heptasaurus_kappelensis.jpg/220px-Heptasaurus_kappelensis.jpg)







{kind=link}
{kind=link}
{kind=link}