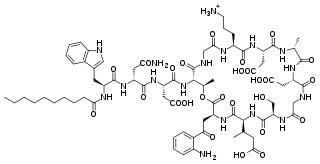
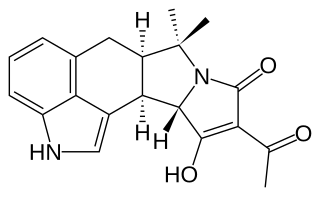
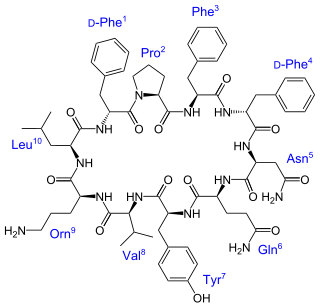
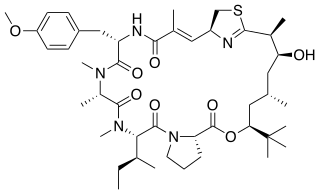
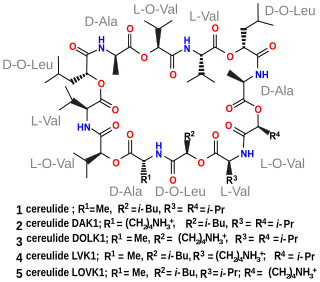
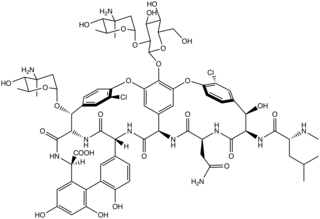
Biosynthesis
Nonribosomal peptides are synthesized by one or more specialized nonribosomal peptide-synthetase (NRPS) enzymes. The NRPS genes for a certain peptide are usually organized in one operon in bacteria and in gene clusters in eukaryotes. However the first fungal NRP to be found was ciclosporin. It is synthesized by a single 1.6MDa NRPS. [4] The enzymes are organized in modules that are responsible for the introduction of one additional amino acid. Each module consists of several domains with defined functions, separated by short spacer regions of about 15 amino acids. [5]
The biosynthesis of nonribosomal peptides shares characteristics with the polyketide and fatty acid biosynthesis. Due to these structural and mechanistic similarities, some nonribosomal peptide synthetases contain polyketide synthase modules for the insertion of acetate or propionate-derived subunits into the peptide chain. [6]
Note that as many as 10% percent of bacterial NRPS are not laid out as large modular proteins, but as separate enzymes. [6] Some NRPS modules deviate from the standard domain structure, and some extra domains have been described. There are also NRPS enzymes that serve as a scaffold for other modifications to the substrate to incorporate unusual amino acids. [7]
Modules
The order of modules and domains of a complete nonribosomal peptide synthetase is as follows:
- Initiation or Starting module: [F/NMT]-A-PCP-
- Elongation or Extending modules: -(C/Cy)-[NMT]-A-PCP-[E]-
- Termination or Releasing module: -(TE/R)
(Order: N-terminus to C-terminus; []: optionally; (): alternatively)
Domains
- F: Formylation (optional)
- A: Adenylation (required in a module)
- PCP: Thiolation and peptide carrier protein with attached 4'-phospho-pantetheine (required in a module)
- C: Condensation forming the amide bond (required in a module)
- Cy: Cyclization into thiazoline or oxazolines (optional)
- Ox: Oxidation of thiazolines or oxazolines to thiazoles or oxazoles (optional)
- Red: Reduction of thiazolines or oxazolines to thiazolidines or oxazolidines (optional)
- E: Epimerization into D-amino acids (optional)
- NMT: N-methylation (optional)
- TE: Termination by a thio-esterase (only found once in a NRPS)
- R: Reduction to terminal aldehyde or alcohol (optional)
- X: Recruits cytochrome P450 enzymes (optional)
Starting stage
- Loading: The first amino acid is activated with ATP as a mixed acyl-phosphoric acid anhydride with AMP by the A-domain and loaded onto the serine-attached 4'-phospho-pantethine (4'PP) sidechain of the PCP-domain catalyzed by the PCP-domain (thiolation).
- Some A domains require interaction with MbtH-like proteins for their activity. [8] [9]
- Sometimes the amino group of the bound amino acid is formylated by an F-domain or methylated by an NMT-domain.
Elongation stages
- Loading: Analogous to the starting stage, each module loads its specific amino acid onto its PCP-domain.
- Condensation: The C-domain catalyzes the amide bond formation between the thioester group of the growing peptide chain from the previous module with the amino group of the current module. The extended peptide is now attached to the current PCP-domain.
- Condensation-Cyclization: Sometimes the C-domain is replaced by a Cy-domain, which, in addition to the amide bond formation, catalyzes the reaction of the serine, threonine, or cysteine sidechain with the amide- N , thereby forming oxazolidines and thiazolidine, respectively.
- Epimerization: Sometimes an E-domain epimerizes the innermost amino acid of the peptide chain into the D-configuration.
- This cycle is repeated for each elongation module.
Termination stage
- Termination: The TE-domain (thio-esterase domain) hydrolyzes the completed polypeptide chain from the PCP-domain of the previous module, thereby often forming cyclic amides (lactams) or cyclic esters (lactones).
- Also, the peptide can be released by an R-domain that reduces the thioester bond to terminal aldehyde or alcohol.
Processing
The final peptide is often modified, e.g., by glycosylation, acylation, halogenation, or hydroxylation. The responsible enzymes are usually associated to the synthetase complex and their genes are organized in the same operons or gene clusters.
Priming and deblocking
To become functional, the 4'-phospho-pantetheine sidechain of acyl-CoA molecules has to be attached to the PCP-domain by 4'PP transferases (Priming) and the S-attached acyl group has to be removed by specialized associated thioesterases (TE-II) (Deblocking).
Substrate specificities
Most domains have a very broad substrate specificity and usually only the A-domain determines which amino acid is incorporated in a module. Ten amino acids that control substrate specificity and can be considered the 'codons' of nonribosomal peptide synthesis have been identified, and rational protein design has yielded methodologies to computationally switch the specificities of A-domains. [10] The condensation C-domain is also believed to have substrate specificity, especially if located behind an epimerase E-domain-containing module where it functions as a 'filter' for the epimerized isomer. Computational methods, such as SANDPUMA [11] and NRPSpredictor2, [12] have been developed to predict substrate specificity from DNA or protein sequence data.
Mixed with polyketides
Due to the similarity with polyketide synthases (PKS), many secondary metabolites are, in fact, fusions of NRPs and polyketides. In essence, this occurs when PK modules follow NRP modules, and vice versa. Although there is high degree of similarity between the Carrier (PCP/ACP) domains of both types of synthetases, the mechanism of condensation is different from a chemical standpoint:
- PKS, carbon-carbon bond formation through Claisen condensation reaction
- NRPs, the C domain catalyzes the amide bond formation between the amino acid it adds to the chain (on the PCP of one module) and the nascent peptide(on the PCP of the next module). [13]