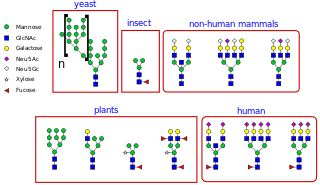
Types
N-linked glycosylation
N-linked glycosylation is a very prevalent form of glycosylation and is important for the folding of many eukaryotic glycoproteins and for cell–cell and cell– extracellular matrix attachment. The N-linked glycosylation process occurs in eukaryotes in the lumen of the endoplasmic reticulum and widely in archaea, but very rarely in bacteria. In addition to their function in protein folding and cellular attachment, the N-linked glycans of a protein can modulate a protein's function, in some cases acting as an on/off switch.
O-linked glycosylation
O-linked glycosylation is a form of glycosylation that occurs in eukaryotes in the Golgi apparatus, [11] but also occurs in archaea and bacteria.
Phosphoserine glycosylation
Xylose, fucose, mannose, and GlcNAc phosphoserine glycans have been reported in the literature. Fucose and GlcNAc have been found only in Dictyostelium discoideum, mannose in Leishmania mexicana , and xylose in Trypanosoma cruzi . Mannose has recently been reported in a vertebrate, the mouse, Mus musculus, on the cell-surface laminin receptor alpha dystroglycan4. It has been suggested this rare finding may be linked to the fact that alpha dystroglycan is highly conserved from lower vertebrates to mammals. [12]
C-mannosylation

A mannose sugar is added to the first tryptophan residue in the sequence W–X–X–W (W indicates tryptophan; X is any amino acid). A C-C bond is formed between the first carbon of the alpha-mannose and the second carbon of the tryptophan. [13] However, not all the sequences that have this pattern are mannosylated. It has been established that, in fact, only two thirds are and that there is a clear preference for the second amino acid to be one of the polar ones (Ser, Ala, Gly and Thr) in order for mannosylation to occur. Recently there has been a breakthrough in the technique of predicting whether or not the sequence will have a mannosylation site that provides an accuracy of 93% opposed to the 67% accuracy if we just consider the WXXW motif. [14]
Thrombospondins are one of the proteins most commonly modified in this way. However, there is another group of proteins that undergo C-mannosylation, type I cytokine receptors. [15] C-mannosylation is unusual because the sugar is linked to a carbon rather than a reactive atom such as nitrogen or oxygen. In 2011, the first crystal structure of a protein containing this type of glycosylation was determined—that of human complement component 8. [16] Currently it is established that 18% of human proteins, secreted and transmembrane undergo the process of C-mannosylation. [14] Numerous studies have shown that this process plays an important role in the secretion of Trombospondin type 1 containing proteins which are retained in the endoplasmic reticulum if they do not undergo C-mannosylation [14] This explains why a type of cytokine receptors, erythropoietin receptor remained in the endoplasmic reticulum if it lacked C-mannosylation sites. [17]
Formation of GPI anchors (glypiation)
Glypiation is a special form of glycosylation that features the formation of a GPI anchor. In this kind of glycosylation a protein is attached to a lipid anchor, via a glycan chain. (See also prenylation.)
Chemical glycosylation
Glycosylation can also be effected using the tools of synthetic organic chemistry. Unlike the biochemical processes, synthetic glycochemistry relies heavily on protecting groups [18] (e.g. the 4,6-O-benzylidene) in order to achieve desired regioselectivity. The other challenge of chemical glycosylation is the stereoselectivity that each glycosidic linkage has two stereo-outcomes, α/β or cis/trans. Generally, the α- or cis-glycoside is more challenging to synthesis. [19] New methods have been developed based on solvent participation or the formation of bicyclic sulfonium ions as chiral-auxiliary groups. [20]
Non-enzymatic glycosylation
The non-enzymatic glycosylation is also known as glycation or non-enzymatic glycation. It is a spontaneous reaction and a type of post-translational modification of proteins meaning it alters their structure and biological activity. It is the covalent attachment between the carbonil group of a reducing sugar (mainly glucose and fructose) and the amino acid side chain of the protein. In this process the intervention of an enzyme is not needed. It takes place across and close to the water channels and the protruding tubules. [21]
At first, the reaction forms temporary molecules which later undergo different reactions (Amadori rearrangements, Schiff base reactions, Maillard reactions, crosslinkings...) and form permanent residues known as Advanced Glycation end-products (AGEs).
AGEs accumulate in long-lived extracellular proteins such as collagen [22] which is the most glycated and structurally abundant protein, especially in humans. Also, some studies have shown lysine may trigger spontaneous non-enzymatic glycosylation. [23]
Role of AGEs
AGEs are responsible for many things. These molecules play an important role especially in nutrition, they are responsible for the brownish color and the aromas and flavors of some foods. It is demonstrated that cooking at high temperature results in various food products having high levels of AGEs. [24]
Having elevated levels of AGEs in the body has a direct impact on the development of many diseases. It has a direct implication in diabetes mellitus type 2 that can lead to many complications such as: cataracts, renal failure, heart damage... [25] And, if they are present at a decreased level, skin elasticity is reduced which is an important symptom of aging. [22]
They are also the precursors of many hormones and regulate and modify their receptor mechanisms at the DNA level. [22]