
Snakes are elongated, limbless, carnivorous reptiles of the suborder Serpentes. Like all other squamates, snakes are ectothermic, amniote vertebrates covered in overlapping scales. Many species of snakes have skulls with several more joints than their lizard ancestors, enabling them to swallow prey much larger than their heads. To accommodate their narrow bodies, snakes' paired organs appear one in front of the other instead of side by side, and most have only one functional lung. Some species retain a pelvic girdle with a pair of vestigial claws on either side of the cloaca. Lizards have independently evolved elongate bodies without limbs or with greatly reduced limbs at least twenty-five times via convergent evolution, leading to many lineages of legless lizards. These resemble snakes, but several common groups of legless lizards have eyelids and external ears, which snakes lack, although this rule is not universal.

Mosasaurs are an extinct group of large aquatic reptiles within the family Mosasauridae that lived during the Late Cretaceous. Their first fossil remains were discovered in a limestone quarry at Maastricht on the Meuse in 1764. They belong to the order Squamata, which includes lizards and snakes.
Tylosaurus is a genus of mosasaur, a large, predatory marine reptile closely related to modern monitor lizards and snakes, from the Late Cretaceous.

Pythonomorpha was originally proposed by paleontologist Edward Drinker Cope (1869) as a reptilian order comprising mosasaurs, which he believed to be close relatives of Ophidia (snakes). The etymology of the term Pythonomorpha comes from the Greek Python and morphe ("form"), and refers to the generally serpentine body plan of members of the group. Cope wrote, "In the mosasauroids, we almost realize the fictions of snake-like dragons and sea-serpents, in which men have been ever prone to indulge. On account of the ophidian part of their affinities, I have called this order Pythonomorpha." Cope incorporated two families, the Clidastidae and the Mosasauridae.

Najash is an extinct genus of basal snake from the Late Cretaceous Candeleros Formation of Patagonia. Like a number of other Cretaceous and living snakes it retained hindlimbs, but Najash is unusual in having well-developed legs that extend outside the rib cage, and a pelvis connected to the spine.
Haasiophis, consisting of the sole species Haasiophis terrasanctus, is an extinct genus of snakes with hind limbs. It is one of three genera of Cenomanian snakes known to have possessed hindlimbs.
Pachyrhachis is an extinct genus of snake with well developed hind legs known from fossils discovered in Ein Yabrud, near Ramallah, in the central West Bank. It is a relatively small snake, measuring more than 1.5 metres (4.9 ft) long at maximum. Pachyrhachis appears to have been an ancient marine snake; the fossils occur in a marine limestone deposit, and the thickened bone of the ribs and vertebrae would have functioned as ballast to decrease the buoyancy of the animal, allowing it to dive beneath the ancient Cretaceous seas that it once inhabited.

Pelvic spurs are external protrusions found around the cloaca in certain superfamilies of snakes belonging to the greater infraorder Alethinophidia. These spurs are made up of the remnants of the femur bone, which is then covered by a corneal spur, or claw-like structure. This femur derives from ancestral hind limbs found in the most recent common ancestor of modern snakes and the other reptiles of the clade Toxicofera, many of which have fully functional front and hind limbs. Due to the fact that the spurs derive from the ancestral state of functional legs, but are no longer functional for locomotion specifically, these structures do meet the criteria for being considered vestigial. Nonetheless, uses for the structures have been thoroughly documented. Species that have external spurs have corresponding muscles, neurological structures, and vascularization to allow for independent movement. The spurs are more pronounced and visible in male specimens and have been observed in use during courtship behavior. The spurs are specifically used in the clasping and stimulation of females by males during courtship and mating. In certain species, males will also use their spurs to engage in combat with one another.
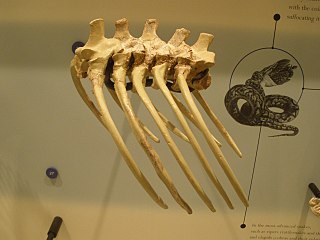
Madtsoiidae is an extinct family of mostly Gondwanan snakes with a fossil record extending from early Cenomanian to late Pleistocene strata located in South America, Africa, India, Australia and Southern Europe. Madtsoiidae include very primitive snakes, which like extant boas and pythons would likely dispatch their prey by constriction. Genera include some of the longest snakes known such as Vasuki, measuring at least 11–15 metres (36–49 ft) long, and the Australian Wonambi and Yurlunggur. As a grouping of basal forms the composition and even the validity of Madtsoiidae is in a state of flux as new pertinent finds are described, with more recent evidence suggesting that it is paraphyletic as previously defined.

Adriosaurus is an extinct genus of squamate which lived in what is now Slovenia and other parts of Europe during the Late Cretaceous. It was small, snake-like reptile, with type species measuring up to 30 cm (12 in) in length. This is the first fossil record of vestigial limbs in lizards. It lost its manus and forearm completely in order to elongate its axial skeleton. These unique anatomical features led to discussions of the evolutionary patterns of limb reduction in Squamata.

Kaganaias is an extinct genus of basal and oldest dolichosaur that lived in what is now Japan during the Early Cretaceous. Kaganaias was semi-aquatic and is the only known aquatic squamate known from before the Cenomanian stage of the Cretaceous. It is also the first to be found in an inland area, instead of on the coast where aquatic squamates are commonly found. Its generic name is derived from Kaga Province, the old name for the Ishikawa Prefecture where the specimens were found, while the species name hakusanensis comes from the mountain that gives its name to Hakusan the city near its find site. The geological formation in which the specimens were found, the Kuwajima Formation, stands alongside the Tetori River and has been the site of numerous other finds including molluscs, dinosaurs, fish, and pterosaurs.

Eupodophis is an extinct genus of snake from the Late Cretaceous period. It has two small hind legs and is considered a transitional form between Cretaceous lizards and limbless snakes. The feature, described as vestigial, was most likely useless to Eupodophis. The type species Eupodophis descouensi was named in 2000 and resides now in the paleontology section of the Mim Museum in Beirut, Lebanon. The specific name is dedicated to the French naturalist Didier Descouens.

Carsosaurus is a genus of extinct amphibious reptiles, in the mosasaur superfamily, containing only the species Carsosaurus marchesetti. It is known from a single individual that lived during the Upper Cretaceous in what is now Slovenia. The specimen is well-preserved, containing many different bones as well as some skin impressions and sternal cartilage. While more remains are needed to be certain, it is generally thought to belong to the Aigialosauridae. In life, it was an amphibious creature that spent most of its time on land, although its later relatives would become fully aquatic.
Parviraptor is a genus of squamate containing one species, Parviraptor estesi, from the Late Jurassic (Tithonian) or Early Cretaceous (Berriasian) Purbeck Limestone Formation of Dorset, England. A second species, Parviraptor gilmorei, was described from the Late Jurassic Morrison Formation of Western North America; it was present in stratigraphic zone 4. However, the second species was subsequently transferred to a separate genus Diablophis. An indeterminate species is known from the Bathonian aged Kirtlington Mammal Bed.
Acteosaurus is an extinct genus of aquatic lizard that lived in the upper Cretaceous period. Its species, A. tommasinii and A. crassicostatus, were described in 1860 and 1993. Though A. crassicostatus is probably a junior synonym for Adriosaurus suessi, A. tommasinii was found to be similar to coniasaurs, mosasauroids, and a sister taxon to modern snakes in 2010.

Tetrapodophis is an extinct genus of lizard from the Early Cretaceous (Aptian) aged Crato Formation of Brazil. It has an elongate snake-like body, with four limbs. Tetrapodophis has been considered by some authors to be one of the oldest members of Ophidia. However, this classification has been disputed by some other authors, who identify Tetrapodophis as a dolichosaurid much less closely related to snakes.

Pachyophis is an extinct genus of Simoliophiidae snakes that were extant during the Cenomanian stage of the Late Cretaceous period. More specifically, it was found to be from the Cenomanian Age about 93.9-100.5 million years ago in the suburb area of Bileca, Herzegovina.

Dolichosauridae is a family of Cretaceous aquatic lizards. They are widely considered to be the earliest and most primitive members of Mosasauria, though some researchers have recovered them as more closely related to snakes.
Ophidiomorpha is a proposed clade composed of snakes and a number of extinct squamate groups. The clade was defined by Placi and Caldwell (2007) as a node-based clade containing the most recent common ancestor of dolichosaurs, adriosaurs, Aphanizocnemus, and fossil and extant Ophidia and all of its descendants.

Mosasauria is a clade of aquatic and semiaquatic squamates that lived during the Cretaceous period. Fossils belonging to the group have been found in all continents around the world. Early mosasaurians like dolichosaurs were small long-bodied lizards that inhabited nearshore coastal and freshwater environments; the Late Cretaceous saw the rise of large marine forms, the mosasaurids, which are the clade's best-known members.

