
Ouranosaurus is a genus of herbivorous basal hadrosauriform dinosaur that lived during the Aptian stage of the Early Cretaceous of modern-day Niger and Cameroon. Ouranosaurus measured about 7 to 8.3 metres long. Two rather complete fossils were found in the Elrhaz Formation, Gadoufaoua deposits, Agadez, Niger, in 1965 and 1970, with a third indeterminate specimen known from the Koum Formation of Cameroon. The animal was named in 1976 by French paleontologist Philippe Taquet; the type species being Ouranosaurus nigeriensis. The generic name is a combination of ourane, a word with multiple meanings, and sauros, the Greek word for lizard. The specific epithet nigeriensis alludes to Niger, its country of discovery. And so, Ouranosaurus nigeriensis could be interpreted as "brave lizard originating from Niger".

Garudimimus is a genus of ornithomimosaur that lived in Asia during the Late Cretaceous. The genus is known from a single specimen found in 1981 by a Soviet-Mongolian paleontological expedition in the Bayan Shireh Formation and formally described in the same year by Rinchen Barsbold; the only species is Garudimimus brevipes. Several interpretations about the anatomical traits of Garudimimus were made in posterior examinations of the specimen, but most of them were criticized during its comprehensive redescription in 2005. Extensive undescribed ornithomimosaur remains at the type locality of Garudimimus may represent additional specimens of the genus.

Huabeisaurus was a genus of dinosaur from the Late Cretaceous. It was a sauropod which lived in what is present-day northern China. The type species, Huabeisaurus allocotus, was first described by Pang Qiqing and Cheng Zhengwu in 2000. Huabeisaurus is known from numerous remains found in the 1990s, which include teeth, partial limbs and vertebrae. Due to its relative completeness, Huabeisaurus represents a significant taxon for understanding sauropod evolution in Asia. Huabeisaurus comes from Kangdailiang and Houyu, Zhaojiagou Town, Tianzhen County, Shanxi province, China. The holotype was found in the unnamed upper member of the Huiquanpu Formation, which is Late Cretaceous (?Cenomanian–?Campanian) in age based on ostracods, charophytes, and fission-track dating.
Haasiophis, consisting of the sole species Haasiophis terrasanctus, is an extinct genus of snakes with hind limbs. It is one of three genera of Cenomanian snakes known to have possessed hindlimbs.

Prognathodon is an extinct genus of marine lizard belonging to the mosasaur family. It is classified as part of the Mosasaurinae subfamily, alongside genera like Mosasaurus and Clidastes. Prognathodon has been recovered from deposits ranging in age from the Campanian to the Maastrichtian in the Middle East, Europe, New Zealand, and North America.

Adriosaurus is an extinct genus of squamate which lived in what is now Slovenia and other parts of Europe during the Late Cretaceous. It was small, snake-like reptile, with type species the measuring up to 30 cm (12 in) in length. This is the first fossil record of vestigial limbs in lizards. It lost its manus and forearm completely in order to elongate its axial skeleton. These unique anatomical features led to discussions of the evolutionary patterns of limb reduction in Squamata.

Anatosuchus is an extinct genus of notosuchian crocodylomorph discovered in Gadoufaoua, Niger, and described by a team of palaeontologists led by the American Paul Sereno in 2003, in the Journal of Vertebrate Paleontology. Its duck-like snout coincidentally makes it resemble a crocoduck, an imagined hybrid animal with the head of a crocodile and the body of a duck.

Thalattosaurus meaning "sea lizard," from the Attic Greek thalatta (θάλαττα), "sea," and sauros (σαῦρος), "lizard," is an extinct genus of marine reptile in the family Thalattosauroidea. They were aquatic diapsids that are known exclusively from the Triassic period. It was a 2–3 metres (6.6–9.8 ft) long shellfish-eating reptile with paddle-like limbs and a down-turned rostrum occurring in the Lower and Middle Triassic Sulphur Mountain Formation of British Columbia as well as the Upper Triassic Hosselkus Limestone of California. It has gained notoriety as a result of studies on general diapsid phylogeny.
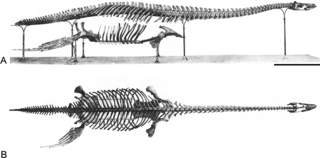
Brancasaurus is a genus of plesiosaur which lived in a freshwater lake in the Early Cretaceous of what is now North Rhine-Westphalia, Germany. With a long neck possessing vertebrae bearing distinctively-shaped "shark fin"-shaped neural spines, and a relatively small and pointed head, Brancasaurus is superficially similar to Elasmosaurus, albeit smaller in size at 3.26 metres (10.7 ft) in length as a subadult.
Hupehsuchia is an order of diapsid reptiles closely related to ichthyosaurs. The group was short-lasting, with a temporal range restricted to the late Olenekian age, spanning only a few million years of the Early Triassic. The order gets its name from Hubei Province, China, from which many specimens have been found. They are probable members of the clade Ichthyosauromorpha.

Cardabiodon is an extinct genus of large mackerel shark that lived about 95 to 91 million years ago (Ma) during the Cenomanian to Turonian of the Late Cretaceous. It is a member of the Cardabiodontidae, a family unique among mackerel sharks due to differing dental structures, and contains the two species C. ricki and C. venator. Cardabiodon fossils have been found in Australia, North America, England, and Kazakhstan. It was likely an antitropical shark that inhabited temperate neritic and offshore oceans between 40° and 60° paleolatitude, similar to the modern porbeagle shark.
Acteosaurus is an extinct genus of aquatic lizard that lived in the upper Cretaceous period. Its species, A. tommasinii and A. crassicostatus, were described in 1860 and 1993. Though A. crassicostatus is probably a junior synonym for Adriosaurus suessi, A. tommasinii was found to be similar to coniasaurs, mosasauroids, and a sister taxon to modern snakes in 2010.

Fodonyx is an extinct genus of rhynchosaur from the middle Triassic epoch of Devon in England. Its fossils were discovered in Otter Sandstone Formation and were first assigned to Rhynchosaurus spenceri. This species was reassigned to its own genus, Fodonyx the holotype of which is EXEMS 60/1985/292, that described by David W. E. Hone and Michael J. Benton in 2008. More recently, one skull was reassigned to the new genus Bentonyx. It is distinguished from other rhynchosaurs by a single autapomorphy, the ventral angling of the paraoccipital processes. In all other rhynchosaurs these processes angle dorsally or are horizontal. It is not known if this conferred any advantage to Fodonyx. Fodonyx was between 40 and 50 cm long.
Dianopachysaurus is an extinct genus of pachypleurosaur known from the lower Middle Triassic of Yunnan Province, southwestern China. It was found in the Middle Triassic Lagerstatte of the Guanling Formation. It was first named by Jun Liu, Olivier Rieppel, Da-Yong Jiang, Jonathan C. Aitchison, Ryosuke Motani, Qi-Yue Zhang, Chang-Yong Zhou and Yuan-Yuan Sun in 2011 and the type species is Dianopachysaurus dingi, thanking a Professor Ding for his help.
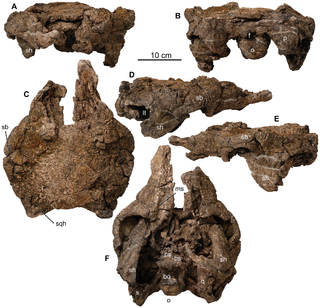
Europelta is a monospecific genus of nodosaurid dinosaur from Spain that lived during the Early Cretaceous in what is now the lower Escucha Formation of the Teruel Province. The type and only species, Europelta carbonensis, is known from two associated partial skeletons, and represents the most complete ankylosaur known from Europe. Europelta was named in 2013 by James I. Kirkland and colleagues. Europelta has an estimated length of 5 metres and weight of 1.3 tonnes, making it the largest member of the clade Struthiosaurini.

Mierasaurus is an extinct genus of sauropod dinosaur from the Early Cretaceous of Utah, United States. The taxon was first described and named in 2017 by Rafael Royo-Torres and colleagues, from a mostly complete skeleton including a disarticulated partial skull and mandible, teeth, multiple vertebrae from along the length of the body, both scapulae, radius and ulna bones, a left manus, a complete pelvis, both femora and the entire left hindlimb. Additionally, they referred a lower jaw and femur from juvenile individuals, which were found nearby, to the genus. Collectively, Mierasaurus is among the most completely known North American sauropods. The genus name honours Bernardo de Miera y Pacheco, the first European scientist to enter what is now Utah. The type species for Mierasaurus is Mierasaurus bobyoungi, named after Robert Glen Young, a paleontologist who researched the Early Cretaceous of Utah.

Mansourasaurus is a genus of herbivorous lithostrotian sauropod dinosaur from the Quseir Formation of Egypt. The type and only species is Mansourasaurus shahinae.

Thalassodraco is an extinct genus of ophthalmosaurid ichthyosaur from the Late Jurassic (Tithonian) Kimmeridge Clay Formation of England. The type species, T. etchesi, was named in 2020, with the epithet in honour of the discoverer of the holotype, Steve Etches.
Barrosasuchus is a genus of peirosaurid notosuchian from the Santonian of Argentina and part of the extensive peirosaurid record of Late Cretaceous Patagonia. It contains one species, Barrosasuchus neuquenianus. B. neuquenianus is known from an almost complete skull and the majority of the articulated postcranial skeleton, making it the best preserved Patagonian peirosaurid.

Monquirasaurus is a monotypic genus of giant pliosaurid from the Cretaceous Paja Formation in Colombia. It contains a single species, M. boyacensis, previously known as Kronosaurus boyacensis.


