Related Research Articles

Staphylococcus aureus is a Gram-positive spherically shaped bacterium, a member of the Bacillota, and is a usual member of the microbiota of the body, frequently found in the upper respiratory tract and on the skin. It is often positive for catalase and nitrate reduction and is a facultative anaerobe that can grow without the need for oxygen. Although S. aureus usually acts as a commensal of the human microbiota, it can also become an opportunistic pathogen, being a common cause of skin infections including abscesses, respiratory infections such as sinusitis, and food poisoning. Pathogenic strains often promote infections by producing virulence factors such as potent protein toxins, and the expression of a cell-surface protein that binds and inactivates antibodies. S. aureus is one of the leading pathogens for deaths associated with antimicrobial resistance and the emergence of antibiotic-resistant strains, such as methicillin-resistant S. aureus (MRSA), is a worldwide problem in clinical medicine. Despite much research and development, no vaccine for S. aureus has been approved.

Methicillin-resistant Staphylococcus aureus (MRSA) is a group of gram-positive bacteria that are genetically distinct from other strains of Staphylococcus aureus. MRSA is responsible for several difficult-to-treat infections in humans. It caused more than 100,000 deaths worldwide attributable to antimicrobial resistance in 2019.

Staphylococcus haemolyticus is a member of the coagulase-negative staphylococci (CoNS). It is part of the skin flora of humans, and its largest populations are usually found at the axillae, perineum, and inguinal areas. S. haemolyticus also colonizes primates and domestic animals. It is a well-known opportunistic pathogen, and is the second-most frequently isolated CoNS. Infections can be localized or systemic, and are often associated with the insertion of medical devices. The highly antibiotic-resistant phenotype and ability to form biofilms make S. haemolyticus a difficult pathogen to treat. Its most closely related species is Staphylococcus borealis.

Staphylococcus epidermidis is a Gram-positive bacterium, and one of over 40 species belonging to the genus Staphylococcus. It is part of the normal human microbiota, typically the skin microbiota, and less commonly the mucosal microbiota and also found in marine sponges. It is a facultative anaerobic bacteria. Although S. epidermidis is not usually pathogenic, patients with compromised immune systems are at risk of developing infection. These infections are generally hospital-acquired. S. epidermidis is a particular concern for people with catheters or other surgical implants because it is known to form biofilms that grow on these devices. Being part of the normal skin microbiota, S. epidermidis is a frequent contaminant of specimens sent to the diagnostic laboratory.

Panton–Valentine leukocidin (PVL) is a cytotoxin—one of the β-pore-forming toxins. The presence of PVL is associated with increased virulence of certain strains (isolates) of Staphylococcus aureus. It is present in the majority of community-associated methicillin-resistant Staphylococcus aureus (CA-MRSA) isolates studied and is the cause of necrotic lesions involving the skin or mucosa, including necrotic hemorrhagic pneumonia. PVL creates pores in the membranes of infected cells. PVL is produced from the genetic material of a bacteriophage that infects Staphylococcus aureus, making it more virulent.
RNAIII is a stable 514 nt regulatory RNA transcribed by the P3 promoter of the Staphylococcus aureus quorum-sensing agr system ). It is the major effector of the agr regulon, which controls the expression of many S. aureus genes encoding exoproteins and cell wall associated proteins plus others encoding regulatory proteins The RNAIII transcript also encodes the 26 amino acid δ-haemolysin peptide (Hld). RNAIII contains many stem loops, most of which match the Shine-Dalgarno sequence involved in translation initiation of the regulated genes. Some of these interactions are inhibitory, others stimulatory; among the former is the regulatory protein Rot. In vitro, RNAIII is expressed post exponentially, inhibiting translation of the surface proteins, notably protein A, while stimulating that of the exoproteins, many of which are tissue-degrading enzymes or cytolysins. Among the latter is the important virulence factor, α-hemolysin (Hla), whose translation RNAIII activates by preventing the formation of an inhibitory foldback loop in the hla mRNA leader.
Lysostaphin is a Staphylococcus simulans metalloendopeptidase. It can function as a bacteriocin (antimicrobial) against Staphylococcus aureus.
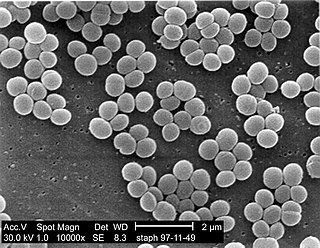
A staphylococcal infection or staph infection is an infection caused by members of the Staphylococcus genus of bacteria.
mecA is a gene found in bacterial cells which allows them to be resistant to antibiotics such as methicillin, penicillin and other penicillin-like antibiotics.
'Staphylococcus aureus delta toxin is a toxin produced by Staphylococcus aureus. It has a wide spectrum of cytolytic activity.

Staphylococcus capitis is a coagulase-negative species (CoNS) of Staphylococcus. It is part of the normal flora of the skin of the human scalp, face, neck, scrotum, and ears and has been associated with prosthetic valve endocarditis, but is rarely associated with native valve infection.

Bullous impetigo is a bacterial skin infection caused by Staphylococcus aureus that results in the formation of large blisters called bullae, usually in areas with skin folds like the armpit, groin, between the fingers or toes, beneath the breast, and between the buttocks. It accounts for 30% of cases of impetigo, the other 70% being non-bullous impetigo.
Staphylococcus is a genus of Gram-positive bacteria in the family Staphylococcaceae from the order Bacillales. Under the microscope, they appear spherical (cocci), and form in grape-like clusters. Staphylococcus species are facultative anaerobic organisms.

Aureolysin is an extracellular metalloprotease expressed by Staphylococcus aureus. This protease is a major contributor to the bacterium's virulence, or ability to cause disease, by cleaving host factors of the innate immune system as well as regulating S. aureus secreted toxins and cell wall proteins. To catalyze its enzymatic activities, aureolysin requires zinc and calcium which it obtains from the extracellular environment within the host.
Staphylococcus schleiferi is a Gram-positive, cocci-shaped bacterium of the family Staphylococcaceae. It is facultatively anaerobic, coagulase-variable, and can be readily cultured on blood agar where the bacterium tends to form opaque, non-pigmented colonies and beta (β) hemolysis. There exists two subspecies under the species S. schleiferi: Staphylococcus schleiferi subsp. schleiferi and Staphylococcus schleiferi subsp. coagulans.
Staphylococcus pseudintermedius is a gram positive coccus bacteria of the genus Staphylococcus found worldwide. It is primarily a pathogen for domestic animals, but has been known to affect humans as well. S. pseudintermedius is an opportunistic pathogen that secretes immune modulating virulence factors, has many adhesion factors, and the potential to create biofilms, all of which help to determine the pathogenicity of the bacterium. Diagnoses of Staphylococcus pseudintermedius have traditionally been made using cytology, plating, and biochemical tests. More recently, molecular technologies like MALDI-TOF, DNA hybridization and PCR have become preferred over biochemical tests for their more rapid and accurate identifications. This includes the identification and diagnosis of antibiotic resistant strains.
The arginine catabolic mobile element (ACME) is a mobile genetic element of Staphylococcus bacterial species. This genetic element provides for several immune modulating functions, including resistance to polyamines which serve as a non-specific immune response both on intact skin and following the inflammatory response in wound healing. Diverse ACME are present in several species of Staphylococcus, including Staphylococcus epidermidis.
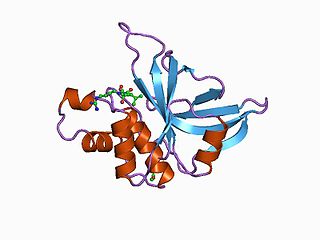
Staphopain A is a secreted cysteine protease produced by Staphylococcus aureus. It was first identified in the S. aureus V8 strain as a papain-like cysteine protease. The protease distinguishes itself from the other major proteases of S. aureus in its very broad specificity and its ability to degrade elastin.
Accessory gene regulator (agr) is a complex 5 gene locus that is a global regulator of virulence in Staphylococcus aureus. It encodes a two-component transcriptional quorum-sensing (QS) system activated by an autoinducing, thiolactone-containing cyclic peptide (AIP).
Kerry L. LaPlante is an American pharmacist, academic and researcher. She is the Dean at the University of Rhode Island College of Pharmacy. She is a Professor of Pharmacy and former department Chair of the Department of Pharmacy Practice at the University of Rhode Island, an Adjunct Professor of Medicine at Brown University, an Infectious Diseases Pharmacotherapy Specialist, and the Director of the Rhode Island Infectious Diseases Fellowship and Research Programs at the Veterans Affairs Medical Center in Providence, Rhode Island.
References
- ↑ Berube, Bryan J.; Sampedro, Georgia R.; Otto, Michael; Bubeck Wardenburg, Juliane (2014-08-01). "The psmα locus regulates production of Staphylococcus aureus alpha-toxin during infection". Infection and Immunity. 82 (8): 3350–3358. doi:10.1128/IAI.00089-14. ISSN 1098-5522. PMC 4136214 . PMID 24866799.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 1. Cheung GYC, Joo HS, Chatterjee SS, Otto M. Phenol-soluble modulins - critical determinants of staphylococcal virulence. FEMS Microbiology Reviews. Blackwell Publishing Ltd; 2014. pp. 698–719. doi:10.1111/1574-6976.12057
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Li S, Huang H, Rao X, Chen W, Wang Z, Hu X. Phenol-soluble modulins: novel virulence-associated peptides of staphylococci REVIEW. Futur Microbiol. 2014;9: 203–216. doi:10.2217/FMB.13.153
- ↑ Jiang Q, Jin Z, Sun B. MgrA negatively regulates biofilm formation and detachment by repressing the expression of psm operons in Staphylococcus aureus. Appl Environ Microbiol. 2018;84: 1–17. doi:10.1128/AEM.001008-18
- ↑ Berube BJ, Sampedro GR, Otto M, Wardenburg JB. The psmα locus regulates production of Staphylococcus aureus alpha-toxin during infection. Infect Immun. 2014;82: 3350–3358. doi:10.1128/IAI.00089-14
- ↑ Yang G, Sau C, Lai W, Cichon J, Li W. Staphylococcus aureus virulent PSMα peptides induce keratinocyte alarmin release to orchestrate IL-17-dependent skin inflammation. 2015;344: 1173–1178. doi:10.1126/science.1249098.Sleep
- ↑ Nguyen TH, Cheung, GYC, Rigby KM, et al. Rapid pathogen-specific recruitment of immune effector cells in the skin by secreted toxins. Nat Microbiol. 2022 Jan;7(1):62-72. doi:10.1038/s41564-021-01012-9
- ↑ Björnsdottir H, Rudin AD, Klose FP, Elmwall J, Welin A, Stylianou M, et al. Phenol-soluble modulin a peptide toxins from aggressive Staphylococcus aureus induce rapid formation of neutrophil extracellular traps through a reactive oxygen species-independent pathway. Front Immunol. 2017;8. doi:10.3389/fimmu.2017.00257
- ↑ Bojer MS, Lindemose S, Vestergaard M, Ingmer H. Quorum sensing-regulated phenol-soluble modulins limit persister cell populations in Staphylococcus aureus. Front. Microbiol. 2018;9. doi:10.3389/fmicb.2018.00255
- ↑ Graves, S. F.; Kobayashi, S. D.; Deleo, F. R. (2010). "Community-associated methicillin-resistant Staphylococcus aureus immune evasion and virulence". Journal of Molecular Medicine. 88 (2): 109–114. doi:10.1007/s00109-009-0573-x. PMC 2852573 . PMID 20049412.
- 1 2 Rasigade, Jean-Philippe; Trouillet-Assant, Sophie; Ferry, Tristan; Diep, Binh An; Sapin, Anaïs; Lhoste, Yannick; Ranfaing, Jérémy; Badiou, Cédric; Benito, Yvonne (2013-01-01). "PSMs of hypervirulent Staphylococcus aureus act as intracellular toxins that kill infected osteoblasts". PLOS ONE. 8 (5): e63176. Bibcode:2013PLoSO...863176R. doi: 10.1371/journal.pone.0063176 . ISSN 1932-6203. PMC 3653922 . PMID 23690994.
- 1 2 3 Kizaki H, Omae Y, Tabuchi F, Saito Y, Sekimizu K, Kaito C. Cell-surface phenol soluble modulins regulate staphylococcus aureus colony spreading. PLoS One. 2016;11: 1–26. doi:10.1371/journal.pone.0164523
- ↑ Schwartz, Kelly; Syed, Adnan K.; Stephenson, Rachel E.; Rickard, Alexander H.; Boles, Blaise R. (2012-06-07). "Functional Amyloids Composed of Phenol Soluble Modulins Stabilize Staphylococcus aureus Biofilms". PLOS Pathogens. 8 (6): e1002744. doi: 10.1371/journal.ppat.1002744 . ISSN 1553-7374. PMC 3369951 . PMID 22685403.