Arabidopsis thaliana, the thale cress, mouse-ear cress or arabidopsis, is a small plant from the mustard family (Brassicaceae), native to Eurasia and Africa. Commonly found along the shoulders of roads and in disturbed land, it is generally considered a weed.

In vascular plants, the roots are the organs of a plant that are modified to provide anchorage for the plant and take in water and nutrients into the plant body, which allows plants to grow taller and faster. They are most often below the surface of the soil, but roots can also be aerial or aerating, that is, growing up above the ground or especially above water.

Auxins are a class of plant hormones with some morphogen-like characteristics. Auxins play a cardinal role in coordination of many growth and behavioral processes in plant life cycles and are essential for plant body development. The Dutch biologist Frits Warmolt Went first described auxins and their role in plant growth in the 1920s. Kenneth V. Thimann became the first to isolate one of these phytohormones and to determine its chemical structure as indole-3-acetic acid (IAA). Went and Thimann co-authored a book on plant hormones, Phytohormones, in 1937.

Phytochromes are a class of photoreceptor proteins found in plants, bacteria and fungi. They respond to light in the red and far-red regions of the visible spectrum and can be classed as either Type I, which are activated by far-red light, or Type II that are activated by red light. Recent advances have suggested that phytochromes also act as temperature sensors, as warmer temperatures enhance their de-activation. All of these factors contribute to the plant's ability to germinate.

Coleoptile is the pointed protective sheath covering the emerging shoot in monocotyledons such as grasses in which few leaf primordia and shoot apex of monocot embryo remain enclosed. The coleoptile protects the first leaf as well as the growing stem in seedlings and eventually, allows the first leaf to emerge. Coleoptiles have two vascular bundles, one on either side. Unlike the flag leaves rolled up within, the pre-emergent coleoptile does not accumulate significant protochlorophyll or carotenoids, and so it is generally very pale. Some preemergent coleoptiles do, however, accumulate purple anthocyanin pigments.

In plant biology, thigmotropism is a directional growth movement which occurs as a mechanosensory response to a touch stimulus. Thigmotropism is typically found in twining plants and tendrils, however plant biologists have also found thigmotropic responses in flowering plants and fungi. This behavior occurs due to unilateral growth inhibition. That is, the growth rate on the side of the stem which is being touched is slower than on the side opposite the touch. The resultant growth pattern is to attach and sometimes curl around the object which is touching the plant. However, flowering plants have also been observed to move or grow their sex organs toward a pollinator that lands on the flower, as in Portulaca grandiflora.

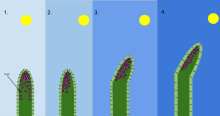
Gravitropism is a coordinated process of differential growth by a plant in response to gravity pulling on it. It also occurs in fungi. Gravity can be either "artificial gravity" or natural gravity. It is a general feature of all higher and many lower plants as well as other organisms. Charles Darwin was one of the first to scientifically document that roots show positive gravitropism and stems show negative gravitropism. That is, roots grow in the direction of gravitational pull and stems grow in the opposite direction. This behavior can be easily demonstrated with any potted plant. When laid onto its side, the growing parts of the stem begin to display negative gravitropism, growing upwards. Herbaceous (non-woody) stems are capable of a degree of actual bending, but most of the redirected movement occurs as a consequence of root or stem growth outside. The mechanism is based on the Cholodny–Went model which was proposed in 1927, and has since been modified. Although the model has been criticized and continues to be refined, it has largely stood the test of time.
In developmental biology, photomorphogenesis is light-mediated development, where plant growth patterns respond to the light spectrum. This is a completely separate process from photosynthesis where light is used as a source of energy. Phytochromes, cryptochromes, and phototropins are photochromic sensory receptors that restrict the photomorphogenic effect of light to the UV-A, UV-B, blue, and red portions of the electromagnetic spectrum.
Shade avoidance is a set of responses that plants display when they are subjected to the shade of another plant. It often includes elongation, altered flowering time, increased apical dominance and altered partitioning of resources. This set of responses is collectively called the shade-avoidance syndrome (SAS).
Cryptochromes are a class of flavoproteins found in plants and animals that are sensitive to blue light. They are involved in the circadian rhythms and the sensing of magnetic fields in a number of species. The name cryptochrome was proposed as a portmanteau combining the chromatic nature of the photoreceptor, and the cryptogamic organisms on which many blue-light studies were carried out.
Phototropins are photoreceptor proteins that mediate phototropism responses in various species of algae, fungi and higher plants. Phototropins can be found throughout the leaves of a plant. Along with cryptochromes and phytochromes they allow plants to respond and alter their growth in response to the light environment. Phototropins may also be important for the opening of stomata and the movement of chloroplasts. These blue light receptors are seen across the entire green plant lineage. When Phototropins are hit with blue light, they induce a signal transduction pathway that alters the plant cells' functions in different ways.
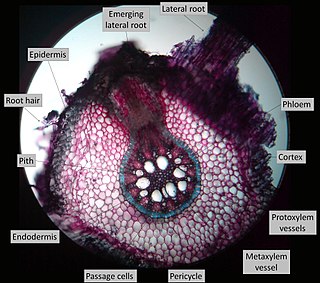
Lateral roots, emerging from the pericycle, extend horizontally from the primary root (radicle) and over time makeup the iconic branching pattern of root systems. They contribute to anchoring the plant securely into the soil, increasing water uptake, and facilitate the extraction of nutrients required for the growth and development of the plant. Lateral roots increase the surface area of a plant's root system and can be found in great abundance in several plant species. In some cases, lateral roots have been found to form symbiotic relationships with rhizobia (bacteria) and mycorrhizae (fungi) found in the soil, to further increase surface area and increase nutrient uptake.

Etiolation is a process in flowering plants grown in partial or complete absence of light. It is characterized by long, weak stems; smaller leaves due to longer internodes; and a pale yellow color (chlorosis). The development of seedlings in the dark is known as "skotomorphogenesis" and leads to etiolated seedlings.
Photoreceptor proteins are light-sensitive proteins involved in the sensing and response to light in a variety of organisms. Some examples are rhodopsin in the photoreceptor cells of the vertebrate retina, phytochrome in plants, and bacteriorhodopsin and bacteriophytochromes in some bacteria. They mediate light responses as varied as visual perception, phototropism and phototaxis, as well as responses to light-dark cycles such as circadian rhythm and other photoperiodisms including control of flowering times in plants and mating seasons in animals.

In botany, the Cholodny–Went model, proposed in 1927, is an early model describing tropism in emerging shoots of monocotyledons, including the tendencies for the shoot to grow towards the light (phototropism) and the roots to grow downward (gravitropism). In both cases the directional growth is considered to be due to asymmetrical distribution of auxin, a plant growth hormone. Although the model has been criticized and continues to be refined, it has largely stood the test of time.

Joanne Chory is an American plant biologist and geneticist. Chory is a professor and director of the Plant Molecular and Cellular Biology Laboratory, at the Salk Institute for Biological Studies and an investigator of the Howard Hughes Medical Institute.
Circadian Clock Associated 1 (CCA1) is a gene that is central to the circadian oscillator of angiosperms. It was first identified in Arabidopsis thaliana in 1993. CCA1 interacts with LHY and TOC1 to form the core of the oscillator system. CCA1 expression peaks at dawn. Loss of CCA1 function leads to a shortened period in the expression of many other genes.
LUX or Phytoclock1 (PCL1) is a gene that codes for LUX ARRHYTHMO, a protein necessary for circadian rhythms in Arabidopsis thaliana. LUX protein associates with Early Flowering 3 (ELF3) and Early Flowering 4 (ELF4) to form the Evening Complex (EC), a core component of the Arabidopsis repressilator model of the plant circadian clock. The LUX protein functions as a transcription factor that negatively regulates Pseudo-Response Regulator 9 (PRR9), a core gene of the Midday Complex, another component of the Arabidopsis repressilator model. LUX is also associated with circadian control of hypocotyl growth factor genes PHYTOCHROME INTERACTING FACTOR 4 (PIF4) and PHYTOCHROME INTERACTING FACTOR 5 (PIF5).
Magnetotropism is the movement or plant growth in response to the stimulus provided by the magnetic field in plants around the world. As a natural environmental factor in the Earth, variations of magnetic field level causes many biological effects, including germination rate, flowering time, photosynthesis, biomass accumulation, activation of cryptochrome, and shoot growth.
EARLY FLOWERING 3 (ELF3) is a plant-specific gene that encodes the hydroxyproline-rich glycoprotein and is required for the function of the circadian clock. ELF3 is one of the three components that make up the Evening Complex (EC) within the plant circadian clock, in which all three components reach peak gene expression and protein levels at dusk. ELF3 serves as a scaffold to bind EARLY FLOWERING 4 (ELF4) and LUX ARRHYTHMO (LUX), two other components of the EC, and functions to control photoperiod sensitivity in plants. ELF3 also plays an important role in temperature and light input within plants for circadian clock entrainment. Additionally, it plays roles in light and temperature signaling that are independent from its role in the EC.