
Basidiomycota is one of two large divisions that, together with the Ascomycota, constitute the subkingdom Dikarya within the kingdom Fungi. Members are known as basidiomycetes. More specifically, Basidiomycota includes these groups: agarics, puffballs, stinkhorns, bracket fungi, other polypores, jelly fungi, boletes, chanterelles, earth stars, smuts, bunts, rusts, mirror yeasts, and Cryptococcus, the human pathogenic yeast.
Chytridiomycota are a division of zoosporic organisms in the kingdom Fungi, informally known as chytrids. The name is derived from the Ancient Greek χυτρίδιον, meaning "little pot", describing the structure containing unreleased zoospores. Chytrids are one of the earliest diverging fungal lineages, and their membership in kingdom Fungi is demonstrated with chitin cell walls, a posterior whiplash flagellum, absorptive nutrition, use of glycogen as an energy storage compound, and synthesis of lysine by the α-amino adipic acid (AAA) pathway.

The Oomycetes, or Oomycota, form a distinct phylogenetic lineage of fungus-like eukaryotic microorganisms within the Stramenopiles. They are filamentous and heterotrophic, and can reproduce both sexually and asexually. Sexual reproduction of an oospore is the result of contact between hyphae of male antheridia and female oogonia; these spores can overwinter and are known as resting spores. Asexual reproduction involves the formation of chlamydospores and sporangia, producing motile zoospores. Oomycetes occupy both saprophytic and pathogenic lifestyles, and include some of the most notorious pathogens of plants, causing devastating diseases such as late blight of potato and sudden oak death. One oomycete, the mycoparasite Pythium oligandrum, is used for biocontrol, attacking plant pathogenic fungi. The oomycetes are also often referred to as water molds, although the water-preferring nature which led to that name is not true of most species, which are terrestrial pathogens.

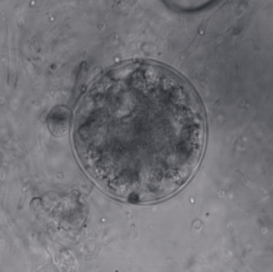
Synchytrium endobioticum is a chytrid fungus that causes the potato wart disease, or black scab. It also infects some other plants of the genus Solanum, though potato is the only cultivated host.
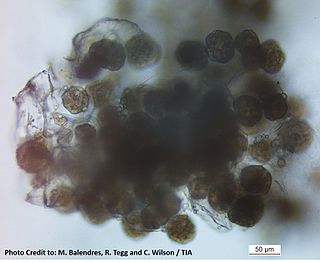
Powdery scab is a disease of potato tubers. It is caused by the cercozoan Spongospora subterranea f. sp. subterranea and is widespread in potato growing countries. Symptoms of powdery scab include small lesions in the early stages of the disease, progressing to raised pustules containing a powdery mass. These can eventually rupture within the tuber periderm. The powdery pustules contain resting spores that release anisokont zoospores to infect the root hairs of potatoes or tomatoes. Powdery scab is a cosmetic defect on tubers, which can result in the rejection of these potatoes. Potatoes which have been infected can be peeled to remove the infected skin and the remaining inside of the potato can be cooked and eaten.
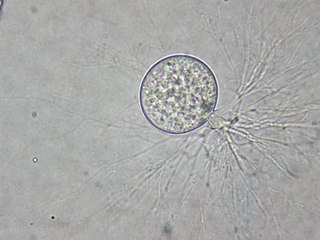
Fungi of the order Chytridiales, like other members of its division, may either have a monocentric thallus or a polycentric rhizomycelium. When the ribosomal genes of members classified in this order were first examined using molecular techniques, it was discovered that the order contained some species that were not related. With the culture and characterization of Chytridium olla, the type species of this order, the limits of the Chytridiales were established. The Chytridiales is now monophyletic and species such as Polychytrium aggregatum, Chytriomyces angularis and Cladochytrium replicatum have been transferred to other orders.

Rhizophydiales are an important group of chytrid fungi. They are found in soil as well as marine and fresh water habitats where they function as parasites and decomposers.

Blastocladiomycota is one of the currently recognized phyla within the kingdom Fungi. Blastocladiomycota was originally the order Blastocladiales within the phylum Chytridiomycota until molecular and zoospore ultrastructural characters were used to demonstrate it was not monophyletic with Chytridiomycota. The order was first erected by Petersen for a single genus, Blastocladia, which was originally considered a member of the oomycetes. Accordingly, members of Blastocladiomycota are often referred to colloquially as "chytrids." However, some feel "chytrid" should refer only to members of Chytridiomycota. Thus, members of Blastocladiomyota are commonly called "blastoclads" by mycologists. Alternatively, members of Blastocladiomycota, Chytridiomycota, and Neocallimastigomycota lumped together as the zoosporic true fungi. Blastocladiomycota contains 5 families and approximately 12 genera. This early diverging branch of kingdom Fungi is the first to exhibit alternation of generations. As well, two (once) popular model organisms—Allomyces macrogynus and Blastocladiella emersonii—belong to this phylum.
Salmacisia is a fungal genus in the family Tilletiaceae. It is a monotypic genus, containing the single species Salmacisia buchloëana, first described as Tilletia buchloëana in 1889, and renamed in 2008. Plants infected by the fungus undergo a phenomenon known as "parasitically induced hermaphroditism", whereby ovary development is induced in otherwise male plants. Because of the pistil-inducing effects of the fungus, the authors have named the species pistil smut; it is the only species in the order Tilletiales known to have hermaphroditic effects.
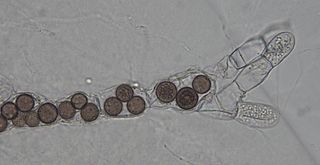
Rozella is a fungal genus of obligate endoparasites of a variety of hosts, including Oomycota, Chytridiomycota, and Blastocladiomycota. Rozella was circumscribed by French mycologist Marie Maxime Cornu in 1872. Considered one of the earliest diverging lineages of fungi, the widespread genus contains 27 species, with the most well studied being Rozella allomycis. Rozella is a member of a large clade of fungi referred to as the Cryptomycota/Rozellomycota. While some can be maintained in dual culture with the host, most have not been cultured, but they have been detected, using molecular techniques, in soil samples, and in freshwater and marine ecosystems. Zoospores have been observed, along with cysts, and the cells of some species are attached to diatoms.

Synchytrium is a large genus of plant pathogens within the phylum Chytridiomycota. Species are commonly known as false rust or wart disease. Approximately 200 species are described, and all are obligate parasites of angiosperms, ferns, or mosses. Early species were mistakenly classified among the higher fungi because of their superficial similarity to the rust fungi. Anton de Bary and Mikhail S. Woronin recognized the true nature of these fungi and established the genus to accommodate Synchytrium taraxaci, which grows on dandelions, and S. succisae, which grows on Succisa pratensis. Synchytrium taraxaci is the type of the genus. The genus has been divided into 6 subgenera based on differences in life cycles.
Olpidium is a fungal genus in the family Olpidiaceae. Members of Olpidium are zoosporic pathogens of plants, animals, fungi, and oomycetes.
Allomyces is a genus of fungi in the family Blastocladiaceae. It was circumscribed by British mycologist Edwin John Butler in 1911. Species in the genus have a polycentric thallus and reproduce sexually or asexually by zoospores that have a whiplash-like flagella. They are mostly isolated from soils in tropical countries, commonly in ponds, rice fields, and slow-moving rivers.
Neocallimastix patriciarum is a species of fungus that lives in the rumen of sheep and other ruminant species. N. patriciarum is an obligate anaerobe and is an important component of the microbial population within the rumen. Only one of a few rumen fungi, this species is interesting and unique within the fungal world. Originally thought to be a flagellate protists, species within the phylum Neocallimastigomycota were first recognized as a fungus by Colin Orpin in 1975 when he demonstrated that they had cell walls of chitin
Chytriomyces is the type genus of fungi in the family Chytriomycetaceae. The genus was described by mycologist John Sidney Karling in 1945. The family, created by Peter Letcher in 2011, contains species with a Group I-type zoospore, distinguishing it from Chytridiaceae members, which have a Group II-type zoospore.
Frederick Kroeber Sparrow was an American mycologist. He was known for his research on aquatic fungi, and in particular the genus Physoderma, and he produced a well-received monograph in 1943 titled The Aquatic Phycomycetes Exclusive of the Saprolegniacea and Pythium; this was republished in 1960 as Aquatic Phycomycetes.
Dendrochytridium is a fungal genus in the order Chytridiales. The genus is monotypic, containing the single saprobic species Dendrochytridium crassum, isolated from detritus collected from an Australian tree canopy. Both the genus and species were described as new to science in 2013. Phylogenetically, Dendrochytridium crassum groups together in a clade with other fungi possessing Group II-type zoospores. These fungi, which include representatives from the genera Chytridium, Phlyctochytrium, Chytriomyces, and Polyphlyctis are classified in the family Chytridiaceae.
Globisporangium sylvaticum is a plant pathogen, an oomycete known to cause root rot and damping off in a multitude of species. These species include apples, carrot, cherry laurel, cress, cucumber, garlic, lettuce, pea, rhododendron, and spinach. Symptoms of infection include stunting, wilt, chlorosis, and browning and eventual necrosis of roots. The pathogen can by identified by the presence of thick, microscopic, round spores within the cells of the root.

Physodermatacae is a family of chytrid fungi in the order Physodermatales. Species in the family have a parasitic relationship with the host's physoderma. This family is distinctive in that it contains a thick wall around the sporangia to resist against unfavorable conditions. Sporangia releases from a host plant when rotting, dispersal is carried through the air. This family is not to be confused or related to basidiomycetes rusts and smut fungi. This parasite is distributed all across the world in aquatic, semi aquatic wetlands and in some ferns.

Aphelidium tribonemae is a species within the Aphelid group. Their classification in the kingdom Fungi is a subject of controversy. Some argue for the classification of aphelids as ‘fungal animals', and for a period of time in the 1950s, aphids were classified as protists due to their amoeboid stage. Recently, molecular phylogenetics placed the aphelids within Opisthosporidia, a super phylum within Opisthokonta. Aphelids have posterior uniflagellate zoospores which place them as Opisthokonts. They are an early diverging lineage in Kingdom Fungi. While the aphelid group only contains three genera, it spans many both freshwater and marine ecosystems.
