Related Research Articles
A restriction enzyme, restriction endonuclease, REase, ENase orrestrictase is an enzyme that cleaves DNA into fragments at or near specific recognition sites within molecules known as restriction sites. Restriction enzymes are one class of the broader endonuclease group of enzymes. Restriction enzymes are commonly classified into five types, which differ in their structure and whether they cut their DNA substrate at their recognition site, or if the recognition and cleavage sites are separate from one another. To cut DNA, all restriction enzymes make two incisions, once through each sugar-phosphate backbone of the DNA double helix.

Neisseria is a large genus of bacteria that colonize the mucosal surfaces of many animals. Of the 11 species that colonize humans, only two are pathogens, N. meningitidis and N. gonorrhoeae.

Salvador Edward Luria was an Italian microbiologist, later a naturalized U.S. citizen. He won the Nobel Prize in Physiology or Medicine in 1969, with Max Delbrück and Alfred Hershey, for their discoveries on the replication mechanism and the genetic structure of viruses. Salvador Luria also showed that bacterial resistance to viruses (phages) is genetically inherited.
In molecular biology, endonucleases are enzymes that cleave the phosphodiester bond within a polynucleotide chain. Some, such as deoxyribonuclease I, cut DNA relatively nonspecifically, while many, typically called restriction endonucleases or restriction enzymes, cleave only at very specific nucleotide sequences. Endonucleases differ from exonucleases, which cleave the ends of recognition sequences instead of the middle (endo) portion. Some enzymes known as "exo-endonucleases", however, are not limited to either nuclease function, displaying qualities that are both endo- and exo-like. Evidence suggests that endonuclease activity experiences a lag compared to exonuclease activity.

Neisseria meningitidis, often referred to as the meningococcus, is a Gram-negative bacterium that can cause meningitis and other forms of meningococcal disease such as meningococcemia, a life-threatening sepsis. The bacterium is referred to as a coccus because it is round, and more specifically a diplococcus because of its tendency to form pairs.

NlaIII is a type II restriction enzyme isolated from Neisseria lactamica. As part of the restriction modification system, NlaIII is able to prevent foreign DNA from integrating into the host genome by cutting double stranded DNA into fragments at specific sequences. This results in further degradation of the fragmented foreign DNA and prevents it from infecting the host genome.
Zinc-finger nucleases (ZFNs) are artificial restriction enzymes generated by fusing a zinc finger DNA-binding domain to a DNA-cleavage domain. Zinc finger domains can be engineered to target specific desired DNA sequences and this enables zinc-finger nucleases to target unique sequences within complex genomes. By taking advantage of endogenous DNA repair machinery, these reagents can be used to precisely alter the genomes of higher organisms. Alongside CRISPR/Cas9 and TALEN, ZFN is a prominent tool in the field of genome editing.
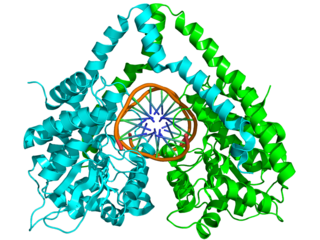
HindIII (pronounced "Hin D Three") is a type II site-specific deoxyribonuclease restriction enzyme isolated from Haemophilus influenzae that cleaves the DNA palindromic sequence AAGCTT in the presence of the cofactor Mg2+ via hydrolysis.

The homing endonucleases are a collection of endonucleases encoded either as freestanding genes within introns, as fusions with host proteins, or as self-splicing inteins. They catalyze the hydrolysis of genomic DNA within the cells that synthesize them, but do so at very few, or even singular, locations. Repair of the hydrolyzed DNA by the host cell frequently results in the gene encoding the homing endonuclease having been copied into the cleavage site, hence the term 'homing' to describe the movement of these genes. Homing endonucleases can thereby transmit their genes horizontally within a host population, increasing their allele frequency at greater than Mendelian rates.
PstI is a type II restriction endonuclease isolated from the Gram negative species, Providencia stuartii.

Molecular cloning is a set of experimental methods in molecular biology that are used to assemble recombinant DNA molecules and to direct their replication within host organisms. The use of the word cloning refers to the fact that the method involves the replication of one molecule to produce a population of cells with identical DNA molecules. Molecular cloning generally uses DNA sequences from two different organisms: the species that is the source of the DNA to be cloned, and the species that will serve as the living host for replication of the recombinant DNA. Molecular cloning methods are central to many contemporary areas of modern biology and medicine.

Genome editing, or genome engineering, or gene editing, is a type of genetic engineering in which DNA is inserted, deleted, modified or replaced in the genome of a living organism. Unlike early genetic engineering techniques that randomly inserts genetic material into a host genome, genome editing targets the insertions to site-specific locations. The basic mechanism involved in genetic manipulations through programmable nucleases is the recognition of target genomic loci and binding of effector DNA-binding domain (DBD), double-strand breaks (DSBs) in target DNA by the restriction endonucleases, and the repair of DSBs through homology-directed recombination (HDR) or non-homologous end joining (NHEJ).

Genetic engineering techniques allow the modification of animal and plant genomes. Techniques have been devised to insert, delete, and modify DNA at multiple levels, ranging from a specific base pair in a specific gene to entire genes. There are a number of steps that are followed before a genetically modified organism (GMO) is created. Genetic engineers must first choose what gene they wish to insert, modify, or delete. The gene must then be isolated and incorporated, along with other genetic elements, into a suitable vector. This vector is then used to insert the gene into the host genome, creating a transgenic or edited organism.

Cas9 is a 160 kilodalton protein which plays a vital role in the immunological defense of certain bacteria against DNA viruses and plasmids, and is heavily utilized in genetic engineering applications. Its main function is to cut DNA and thereby alter a cell's genome. The CRISPR-Cas9 genome editing technique was a significant contributor to the Nobel Prize in Chemistry in 2020 being awarded to Emmanuelle Charpentier and Jennifer Doudna.

Daisy Roulland-Dussoix was a Swiss molecular microbiologist. She was one of the discoverers of restriction enzymes during her doctoral studies. There is controversy over whether she should have received the 1978 Nobel prize in Physiology and Medicine, which was awarded to Hamilton O. Smith, Daniel Nathans, and Werner Arber.

In bacteria, phasevarions mediate a coordinated change in the expression of multiple genes or proteins. This occurs via phase variation of a single DNA methyltransferase. Phase variation of methyltransferase expression results in differential methylation throughout the bacterial genome, leading to variable expression of multiple genes through epigenetic mechanisms.
Since antiretroviral therapy requires a lifelong treatment regimen, research to find more permanent cures for HIV infection is currently underway. It is possible to synthesize zinc finger nucleotides with zinc finger components that selectively bind to specific portions of DNA. Conceptually, targeting and editing could focus on host cellular co-receptors for HIV or on proviral HIV DNA.
References
- ↑ Luria SE, Human ML (1952). "A nonhereditary, host-induced variation of bacterial viruses". J. Bacteriol. 64 (4): 557–69. doi:10.1128/JB.64.4.557-569.1952. PMC 169391 . PMID 12999684.
- ↑ Luria SE (1953). "Host-induced modifications of viruses". Cold Spring Harb. Symp. Quant. Biol. 18: 237–44. doi:10.1101/sqb.1953.018.01.034. PMID 13168990.
- 1 2 Salvator E Luria. A Slot Machine, A Broken Test Tube: An Autobiography. Harper & Row, New York: 1984. Pp. 228. ISBN 0-06-015260-5 (USA and Canada)
- ↑ BERTANI G, WEIGLE JJ (1953). "Host controlled variation in bacterial viruses". J. Bacteriol. 65 (2): 113–21. doi:10.1128/JB.65.2.113-121.1953. PMC 169650 . PMID 13034700.
- ↑ DUSSOIX D, ARBER W (1962). "Host specificity of DNA produced by Escherichia coli. II. Control over acceptance of DNA from infecting phage lambda". J. Mol. Biol. 5: 37–49. doi:10.1016/S0022-2836(62)80059-X. PMID 13888713.
- ↑ Nathans D, Smith HO (1975). "Restriction endonucleases in the analysis and restructuring of dna molecules". Annu. Rev. Biochem. 44: 273–93. doi:10.1146/annurev.bi.44.070175.001421. PMID 166604.
- ↑ Rodic, A; Blagojevic, B; Zdobnov, E; Djordjevic, M; Djordjevic, M (24 February 2017). "Understanding key features of bacterial restriction-modification systems through quantitative modeling". BMC systems biology. 11 (Suppl 1): 377. doi:10.1186/s12918-016-0377-x. PMID 28466789.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ↑ Wilson G (1991). "Organization of Restriction-Modification Systems". Nucleic Acids Research . 19 (10): 2539–2566. doi:10.1093/nar/19.10.2539. PMC 328170 . PMID 2041731.
- ↑ Wilson G (1991). "Restriction and Modification Systems". Annual Review of Genetics. 25: 585–627. doi:10.1146/annurev.ge.25.120191.003101. PMID 1812816.
- ↑ Loenen WA (2013). "The other face of restriction: modification-dependent enzymes". Nucleic Acids Research. 42 (1): 56–69. doi:10.1093/nar/gkt747. PMC 3874153 . PMID 23990325.
- 1 2 3 Budroni S, Siena E, Dunning Hotopp JC, Seib KL, Serruto D, Nofroni C, Comanducci M, Riley DR, Daugherty SC, Angiuoli SV, Covacci A, Pizza M, Rappuoli R, Moxon ER, Tettelin H, Medini D (2011). "Neisseria meningitidis is structured in clades associated with restriction modification systems that modulate homologous recombination". Proc. Natl. Acad. Sci. U.S.A. 108 (11): 4494–9. Bibcode:2011PNAS..108.4494B. doi: 10.1073/pnas.1019751108 . PMC 3060241 . PMID 21368196.
- ↑ Claus H, Friedrich A, Frosch M, Vogel U (2000). "Differential distribution of novel restriction-modification systems in clonal lineages of Neisseria meningitidis". J. Bacteriol. 182 (5): 1296–303. doi:10.1128/jb.182.5.1296-1303.2000. PMC 94415 . PMID 10671450.
- ↑ Ambur OH, Frye SA, Nilsen M, Hovland E, Tønjum T (2012). "Restriction and sequence alterations affect DNA uptake sequence-dependent transformation in Neisseria meningitidis". PLOS ONE. 7 (7): e39742. Bibcode:2012PLoSO...739742A. doi: 10.1371/journal.pone.0039742 . PMC 3388099 . PMID 22768309.
- ↑ Caugant DA, Maiden MC (2009). "Meningococcal carriage and disease--population biology and evolution". Vaccine. 27 Suppl 2 (4): B64–70. doi:10.1016/j.vaccine.2009.04.061. PMC 2719693 . PMID 19464092.
- ↑ Kusano K (1995). "Restriction-Modification Systems as Genomic Parasites in Competition for Specific Sequences". Proceedings of the National Academy of Sciences of the United States of America. 92 (24): 11095–11099. Bibcode:1995PNAS...9211095K. doi: 10.1073/pnas.92.24.11095 . PMC 40578 . PMID 7479944.
- ↑ Oliveira, Pedro H.; Touchon, Marie; Rocha, Eduardo P.C. (2016-05-17). "Regulation of genetic flux between bacteria by restriction–modification systems". Proceedings of the National Academy of Sciences. 113 (20): 5658–5663. Bibcode:2016PNAS..113.5658O. doi: 10.1073/pnas.1603257113 . ISSN 0027-8424. PMC 4878467 . PMID 27140615.
- ↑ Wayengera M (2003). "HIV and Gene Therapy: The proposed [R-M enzymatic] model for a gene therapy against HIV". Makerere Med J. 38: 28–30.
- ↑ Wayengera M, Kajumbula H, Byarugaba W (2007). "Frequency and site mapping of HIV-1/SIVcpz, HIV-2/SIVsmm and Other SIV gene sequence cleavage by various bacteria restriction enzymes: Precursors for a novel HIV inhibitory product". Afr J Biotechnol. 6 (10): 1225–1232.
- ↑ Schiffer JT, Aubert M, Weber ND, Mintzer E, Stone D, Jerome KR (2012). "Targeted DNA mutagenesis for the cure of chronic viral infections". Journal of Virology. 86 (17): 8920–36. doi:10.1128/JVI.00052-12. PMC 3416169 . PMID 22718830.
- ↑ Manjunath N, Yi G, Dang Y, Shankar P (2013). "Newer gene editing technologies toward HIV gene therapy". Viruses. 5 (11): 2748–66. doi: 10.3390/v5112748 . PMC 3856413 . PMID 24284874.
- ↑ Stetson DB, Ko JS, Heidmann T, Medzhitov R (2008). "Trex1 prevents cell intrinsic initiation of autoimmunity". Cell. 134 (4): 587–598. doi:10.1016/j.cell.2008.06.032. PMC 2626626 . PMID 18724932.
- ↑ Gasior SL, Roy-Engel AM, Deininger PL (2008). "ERCC1/XPF limits L1 retrotransposition". DNA Repair. 7 (6): 983–989. doi:10.1016/j.dnarep.2008.02.006. PMC 2483505 . PMID 18396111.
- ↑ Kim YG, Cha J, Chandrasegaran S (February 1996). "Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain". Proc. Natl. Acad. Sci. U.S.A. 93 (3): 1156–60. Bibcode:1996PNAS...93.1156K. doi: 10.1073/pnas.93.3.1156 . PMC 40048 . PMID 8577732.
- ↑ Tebas P, Stein D, Tang WW, Frank I, Wang SQ, Lee G, et al. (2014). "Gene editing of CCR5 in autologous CD4 T cells of persons infected with HIV". N Engl J Med. 370 (10): 901–910. doi:10.1056/NEJMoa1300662. PMC 4084652 . PMID 24597865.
- 1 2 Oliveira, PH; Touchon, M; Rocha, EPC (2014). "The interplay of restriction-modification systems with mobile genetic elements and their prokaryotic hosts". Nucleic Acids Res. 42 (16): 10618–10631. doi:10.1093/nar/gku734. PMC 4176335 . PMID 25120263.