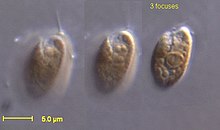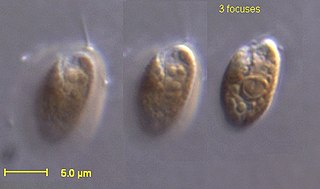
The cryptomonads are a group of algae, most of which have plastids. They are common in freshwater, and also occur in marine and brackish habitats. Each cell is around 10–50 μm in size and flattened in shape, with an anterior groove or pocket. At the edge of the pocket there are typically two slightly unequal flagella.

Euglena is a genus of single cell flagellate eukaryotes. It is the best known and most widely studied member of the class Euglenoidea, a diverse group containing some 54 genera and at least 200 species. Species of Euglena are found in fresh water and salt water. They are often abundant in quiet inland waters where they may bloom in numbers sufficient to color the surface of ponds and ditches green (E. viridis) or red (E. sanguinea).

Chlamydomonas is a genus of green algae consisting of about 150 species of unicellular flagellates, found in stagnant water and on damp soil, in freshwater, seawater, and even in snow as "snow algae". Chlamydomonas is used as a model organism for molecular biology, especially studies of flagellar motility and chloroplast dynamics, biogenesis, and genetics. One of the many striking features of Chlamydomonas is that it contains ion channels (channelrhodopsins) that are directly activated by light. Some regulatory systems of Chlamydomonas are more complex than their homologs in Gymnosperms, with evolutionarily related regulatory proteins being larger and containing additional domains.
Cryptomonas is the name-giving genus of the Cryptomonads established by German biologist Christian Gottfried Ehrenberg in 1831. The algae are common in freshwater habitats and brackish water worldwide and often form blooms in greater depths of lakes. The cells are usually brownish or greenish in color and are characteristic of having a slit-like furrow at the anterior. They are not known to produce any toxins. They are used to feed small zooplankton, which is the food source for small fish in fish farms. Many species of Cryptomonas can only be identified by DNA sequencing. Cryptomonas can be found in several marine ecosystems in Australia and South Korea.
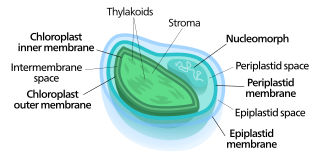
Nucleomorphs are small, vestigial eukaryotic nuclei found between the inner and outer pairs of membranes in certain plastids. They are thought to be vestiges of primitive red and green algal nuclei that were engulfed by a larger eukaryote. Because the nucleomorph lies between two sets of membranes, nucleomorphs support the endosymbiotic theory and are evidence that the plastids containing them are complex plastids. Having two sets of membranes indicate that the plastid, a prokaryote, was engulfed by a eukaryote, an alga, which was then engulfed by another eukaryote, the host cell, making the plastid an example of secondary endosymbiosis.

Tetraselmis is a genus of phytoplankton. Tetraselmis is a green algal genus within the order Chlorodendrales, and they are characterized by their intensely-colored green chloroplast, their flagellated cell bodies, the presence of a pyrenoid within the chloroplast, and a scale-produced thecal-wall. Species within this genus are found in both marine and freshwater ecosystems across the globe; their habitat range is mainly limited by water depth due to their photosynthetic nature. Thus, they live in diverse water environments if enough nutrients and light are available for net photosynthetic activity. Tetraselmis species have proven to be useful for both research and industry. Tetraselmis species have been studied for understanding plankton growth rates, and recently a colonial species is being used to gain an understanding of multicellularity evolution. Additionally, many species are currently being examined for their use as biofuels due to their high lipid content.
The cryptophyceae are a class of algae, most of which have plastids. About 220 species are known, and they are common in freshwater, and also occur in marine and brackish habitats. Each cell is around 10–50 μm in size and flattened in shape, with an anterior groove or pocket. At the edge of the pocket there are typically two slightly unequal flagella.

The raphidophytes, formally known as Raphidophycidae or Raphidophyceae, are a small group of eukaryotic algae that includes both marine and freshwater species. All raphidophytes are unicellular, with large cells, but no cell walls. Raphidophytes possess a pair of flagella, organised such that both originate from the same invagination. One flagellum points forwards, and is covered in hair-like mastigonemes, while the other points backwards across the cell surface, lying within a ventral groove. Raphidophytes contain numerous ellipsoid chloroplasts, which contain chlorophylls a, c1 and c2. They also make use of accessory pigments including β-carotene and diadinoxanthin. Unlike other heterokontophytes, raphidophytes do not possess the photoreceptive organelle typical of this group.
Nephroselmis is a genus of green algae. It has been placed in the family Nephroselmidaceae, although a 2009 study suggests that it should be separated into its own class, Nephroselmidophyceae. One species can be an endosymbiont of Hatena arenicola.
Cryptomonada is a proposed super-class of Cryptists linking Cryptophyceae and Goniomonadea, which was first proposed by Cavalier-Smith in 2004. He made this proposal based on three shared morphological characteristics: Presence of a periplast, ejectisomes with secondary scroll, and mitochondrial cristae with flat tubules.
Goniomonas is a genus of Cryptomonads and contains five species. It is a genus of single-celled eukaryotes, including both freshwater and marine species. It lacks plastids, which is very unusual among all of the Cryptophyte genera. It may reflect one of only a small number of times that the Cryptophytes evolved into freshwater habitats. Goniomonas seems to have a number of freshwater relatives which have not yet been cultured and named.
Geminigera /ˌdʒɛmɪnɪˈdʒɛɹə/ is a genus of cryptophyte from the family Geminigeraceae. Named for its unique pyrenoids, Geminigera is a genus with a single mixotrophic species. It was discovered in 1968 and is known for living in very cold temperatures such as under the Antarctic ice. While originally considered to be part of the genus Cryptomonas, the genus Geminigera was officially described in 1991 by D. R. A. Hill.

Guillardia is a genus of marine biflagellate cryptomonad algae with a plastid obtained through secondary endosymbiosis of a red alga.

Hemiselmis is a genus of cryptomonads.
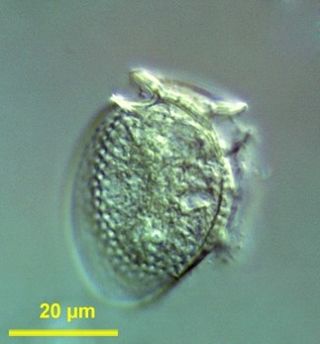
Dinophysis is a genus of dinoflagellates common in tropical, temperate, coastal and oceanic waters. It was first described in 1839 by Christian Gottfried Ehrenberg.
Geminigeraceae is a family of cryptophytes containing the five genera Geminigera, Guillardia, Hanusia, Proteomonas and Teleaulax. They are characterised by chloroplasts containing Cr-phycoerythrin 545, and an inner periplast component (IPC) comprising "a sheet or a sheet and multiple plates if diplomorphic". The nucleomorphs are never in the pyrenoid, and there is never a scalariform furrow. The cells do, however, have a long, keeled rhizostyle with lamellae (wings).
Pyrenomonadaceae is a family of cryptomonads which includes three or four known genera. They are distinguished from other cryptomonads by their nucleomorphs being imbedded into the pyrenoid, and the presence of distinctive pigment phycoerythrin 545.
Chroomonadaceae is a family of cryptomonads first recognized by Clay et al in 1999 as including genera Chroomonas, Falcomonas, and Komma. Following a molecular phylogenic study in 2002, Hemiselmis was also placed within the Chroomonadaceae. Today, the family is generally recognized as sister to the Pyrenomonadaceae.
Cryptochrysis is a formerly recognized genus of cryptomonads first proposed by Adolf Pascher in 1911. He initially treated it as the sole genus in family Cryptochrysidaceae, but later treated it as a member of the Cryptochrysideae subfamily of Cryptomonadaceae, along with Rhodomonas, Chroomonas, and Cyanomonas. In 1967, R.W. Butcher relegated the group to a subgenus within Chroomonas.
Rhinomonas fulva is a phytoplanktonic species of cryptomonad given its current designation in 1988.