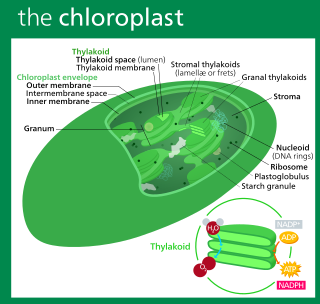
A chloroplast is a type of membrane-bound organelle known as a plastid that conducts photosynthesis mostly in plant and algal cells. The photosynthetic pigment chlorophyll captures the energy from sunlight, converts it, and stores it in the energy-storage molecules ATP and NADPH while freeing oxygen from water in the cells. The ATP and NADPH is then used to make organic molecules from carbon dioxide in a process known as the Calvin cycle. Chloroplasts carry out a number of other functions, including fatty acid synthesis, amino acid synthesis, and the immune response in plants. The number of chloroplasts per cell varies from one, in unicellular algae, up to 100 in plants like Arabidopsis and wheat.
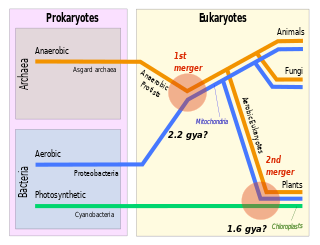
Symbiogenesis is the leading evolutionary theory of the origin of eukaryotic cells from prokaryotic organisms. The theory holds that mitochondria, plastids such as chloroplasts, and possibly other organelles of eukaryotic cells are descended from formerly free-living prokaryotes taken one inside the other in endosymbiosis. Mitochondria appear to be phylogenetically related to Rickettsiales bacteria, while chloroplasts are thought to be related to cyanobacteria.
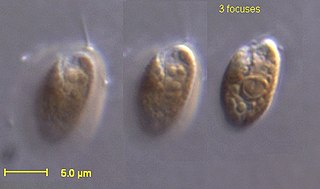
The cryptomonads are a group of algae, most of which have plastids. They are common in freshwater, and also occur in marine and brackish habitats. Each cell is around 10–50 μm in size and flattened in shape, with an anterior groove or pocket. At the edge of the pocket there are typically two slightly unequal flagella.
The plastid is a membrane-bound organelle found in the cells of plants, algae, and some other eukaryotic organisms. They are considered to be intracellular endosymbiotic cyanobacteria. Examples include chloroplasts, chromoplasts, and leucoplasts.

The chlorarachniophytes are a small group of exclusively marine algae widely distributed in tropical and temperate waters. They are typically mixotrophic, ingesting bacteria and smaller protists as well as conducting photosynthesis. Normally they have the form of small amoebae, with branching cytoplasmic extensions that capture prey and connect the cells together, forming a net. They may also form flagellate zoospores, which characteristically have a single subapical flagellum that spirals backwards around the cell body, and walled coccoid cells.

Chromista is a proposed but polyphyletic biological kingdom consisting of single-celled and multicellular eukaryotic species that share similar features in their photosynthetic organelles (plastids). It includes all eukaryotes whose plastids contain chlorophyll c and are surrounded by four membranes. If the ancestor already possessed chloroplasts derived by endosymbiosis from red algae, all non-photosynthetic Chromista have secondarily lost the ability to photosynthesise. Its members might have arisen independently as separate evolutionary groups from the last eukaryotic common ancestor.
Cryptomonas is the name-giving genus of the Cryptomonads established by German biologist Christian Gottfried Ehrenberg in 1831. The algae are common in freshwater habitats and brackish water worldwide and often form blooms in greater depths of lakes. The cells are usually brownish or greenish in color and are characteristic of having a slit-like furrow at the anterior. They are not known to produce any toxins. They are used to feed small zooplankton, which is the food source for small fish in fish farms. Many species of Cryptomonas can only be identified by DNA sequencing. Cryptomonas can be found in several marine ecosystems in Australia and South Korea.
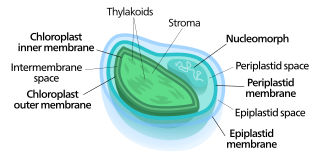
Nucleomorphs are small, vestigial eukaryotic nuclei found between the inner and outer pairs of membranes in certain plastids. They are thought to be vestiges of primitive red and green algal nuclei that were engulfed by a larger eukaryote. Because the nucleomorph lies between two sets of membranes, nucleomorphs support the endosymbiotic theory and are evidence that the plastids containing them are complex plastids. Having two sets of membranes indicate that the plastid, a prokaryote, was engulfed by a eukaryote, an alga, which was then engulfed by another eukaryote, the host cell, making the plastid an example of secondary endosymbiosis.

Kleptoplasty or kleptoplastidy is a process in symbiotic relationships whereby plastids, notably chloroplasts from algae, are sequestered by the host. The word is derived from Kleptes (κλέπτης) which is Greek for thief. The alga is eaten normally and partially digested, leaving the plastid intact. The plastids are maintained within the host, temporarily continuing photosynthesis and benefiting the host.

The Archaeplastida are a major group of eukaryotes, comprising the photoautotrophic red algae (Rhodophyta), green algae, land plants, and the minor group glaucophytes. It also includes the non-photosynthetic lineage Rhodelphidia, a predatorial (eukaryotrophic) flagellate that is sister to the Rhodophyta, and probably the microscopic picozoans. The Archaeplastida have chloroplasts that are surrounded by two membranes, suggesting that they were acquired directly through a single endosymbiosis event by feeding on a cyanobacterium. All other groups which have chloroplasts, besides the amoeboid genus Paulinella, have chloroplasts surrounded by three or four membranes, suggesting they were acquired secondarily from red or green algae. Unlike red and green algae, glaucophytes have never been involved in secondary endosymbiosis events.
The cryptophyceae are a class of algae, most of which have plastids. About 220 species are known, and they are common in freshwater, and also occur in marine and brackish habitats. Each cell is around 10–50 μm in size and flattened in shape, with an anterior groove or pocket. At the edge of the pocket there are typically two slightly unequal flagella.
An apicoplast is a derived non-photosynthetic plastid found in most Apicomplexa, including Toxoplasma gondii, and Plasmodium falciparum and other Plasmodium spp., but not in others such as Cryptosporidium. It originated from algae through secondary endosymbiosis; there is debate as to whether this was a green or red alga. The apicoplast is surrounded by four membranes within the outermost part of the endomembrane system. The apicoplast hosts important metabolic pathways like fatty acid synthesis, isoprenoid precursor synthesis and parts of the heme biosynthetic pathway.

Red algae, or Rhodophyta, are one of the oldest groups of eukaryotic algae. The Rhodophyta comprises one of the largest phyla of algae, containing over 7,000 currently recognized species with taxonomic revisions ongoing. The majority of species (6,793) are found in the Florideophyceae (class), and mostly consist of multicellular, marine algae, including many notable seaweeds. Red algae are abundant in marine habitats but relatively rare in freshwaters. Approximately 5% of red algae species occur in freshwater environments, with greater concentrations found in warmer areas. Except for two coastal cave dwelling species in the asexual class Cyanidiophyceae, there are no terrestrial species, which may be due to an evolutionary bottleneck in which the last common ancestor lost about 25% of its core genes and much of its evolutionary plasticity.
Rhodomonas is a genus of cryptomonads. It is characterized by its red colour, the square-shaped plates of its inner periplast, its short furrow ending in a gullet, and a distinctly shaped chloroplast closely associated with its nucleomorph. Historically, Rhodomonas was characterized by its red chloroplast alone, but this no longer occurs as its taxonomy has become increasingly based on molecular and cellular data. Currently, there is some debate about the taxonomic validity of Rhodomonas as a genus and further research is needed to verify its taxonomic status. Rhodomonas is typically found in marine environments, although freshwater reports exist. It is commonly used as a live feed for various aquaculture species.

Hemiselmis is a genus of cryptomonads.
Dinophysis is a genus of dinoflagellates common in tropical, temperate, coastal and oceanic waters. It was first described in 1839 by Christian Gottfried Ehrenberg.
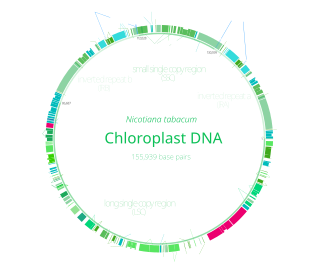
Chloroplast DNA (cpDNA) is the DNA located in chloroplasts, which are photosynthetic organelles located within the cells of some eukaryotic organisms. Chloroplasts, like other types of plastid, contain a genome separate from that in the cell nucleus. The existence of chloroplast DNA was identified biochemically in 1959, and confirmed by electron microscopy in 1962. The discoveries that the chloroplast contains ribosomes and performs protein synthesis revealed that the chloroplast is genetically semi-autonomous. The first complete chloroplast genome sequences were published in 1986, Nicotiana tabacum (tobacco) by Sugiura and colleagues and Marchantia polymorpha (liverwort) by Ozeki et al. Since then, a great number of chloroplast DNAs from various species have been sequenced.
A plastid is a membrane-bound organelle found in plants, algae and other eukaryotic organisms that contribute to the production of pigment molecules. Most plastids are photosynthetic, thus leading to color production and energy storage or production. There are many types of plastids in plants alone, but all plastids can be separated based on the number of times they have undergone endosymbiotic events. Currently there are three types of plastids; primary, secondary and tertiary. Endosymbiosis is reputed to have led to the evolution of eukaryotic organisms today, although the timeline is highly debated.
Lepidodinium is a genus of dinoflagellates belonging to the family Gymnodiniaceae.
