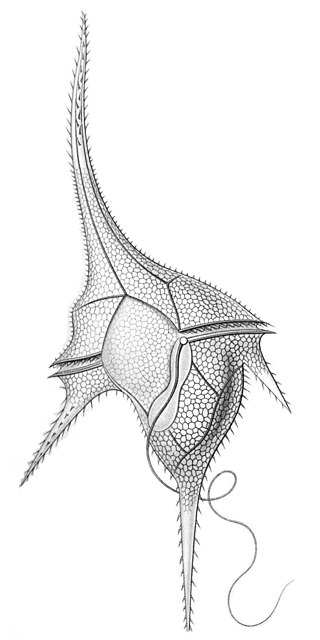
The dinoflagellates are a monophyletic group of single-celled eukaryotes constituting the phylum Dinoflagellata and are usually considered protists. Dinoflagellates are mostly marine plankton, but they also are common in freshwater habitats. Their populations vary with sea surface temperature, salinity, and depth. Many dinoflagellates are photosynthetic, but a large fraction of these are in fact mixotrophic, combining photosynthesis with ingestion of prey.
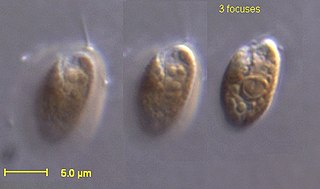
The cryptomonads are a group of algae, most of which have plastids. They are common in freshwater, and also occur in marine and brackish habitats. Each cell is around 10–50 μm in size and flattened in shape, with an anterior groove or pocket. At the edge of the pocket there are typically two slightly unequal flagella.

Chromista is a proposed but polyphyletic biological kingdom, refined from the Chromalveolata, consisting of single-celled and multicellular eukaryotic species that share similar features in their photosynthetic organelles (plastids). It includes all eukaryotes whose plastids contain chlorophyll c and are surrounded by four membranes. If the ancestor already possessed chloroplasts derived by endosymbiosis from red algae, all non-photosynthetic Chromista have secondarily lost the ability to photosynthesise. Its members might have arisen independently as separate evolutionary groups from the last eukaryotic common ancestor.

Kleptoplasty or kleptoplastidy is a process in symbiotic relationships whereby plastids, notably chloroplasts from algae, are sequestered by the host. The word is derived from Kleptes (κλέπτης) which is Greek for thief. The alga is eaten normally and partially digested, leaving the plastid intact. The plastids are maintained within the host, temporarily continuing photosynthesis and benefiting the host.
Chromalveolata was a eukaryote supergroup present in a major classification of 2005, then regarded as one of the six major groups within the eukaryotes. It was a refinement of the kingdom Chromista, first proposed by Thomas Cavalier-Smith in 1981. Chromalveolata was proposed to represent the organisms descended from a single secondary endosymbiosis involving a red alga and a bikont. The plastids in these organisms are those that contain chlorophyll c.

Paulinella is a genus of at least eleven species including both freshwater and marine amoeboids. Like many members of euglyphids it is covered by rows of siliceous scales, and use filose pseudopods to crawl over the substrate of the benthic zone.

Protozoa are a polyphyletic group of single-celled eukaryotes, either free-living or parasitic, that feed on organic matter such as other microorganisms or organic debris. Historically, protozoans were regarded as "one-celled animals".

Guillardia is a genus of marine biflagellate cryptomonad algae with a plastid obtained through secondary endosymbiosis of a red alga.

Hemiselmis is a genus of cryptomonads.
Amoebophyra is a genus of dinoflagellates. Amoebophyra is a syndinian parasite that infects free-living dinoflagellates that are attributed to a single species by using several host-specific parasites. It acts as "biological control agents for red tides and in defining species of Amoebophrya." Researchers have found a correlation between a large amount of host specify and the impact host parasites may have on other organisms. Due to the host specificity found in each strain of Amoebophrya's physical makeup, further studies need to be tested to determine whether the Amoebophrya can act as a control against harmful algal blooms.
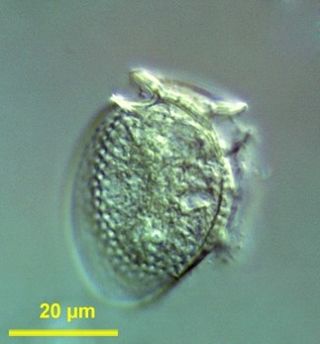
Dinophysis is a genus of dinoflagellates common in tropical, temperate, coastal and oceanic waters. It was first described in 1839 by Christian Gottfried Ehrenberg.

Raphidiophrys contractilis is a species of freshwater centrohelid.
Mesodinium chamaeleon is a ciliate of the genus Mesodinium. It is known for being able to consume and maintain algae endosymbiotically for days before digesting the algae. It has the ability to eat red and green algae, and afterwards using the chlorophyll granules from the algae to generate energy, turning itself from being a heterotroph into an autotroph. The species was discovered in January 2012 outside the coast of Nivå, Denmark by professor Øjvind Moestrup.
Vahlkampfia avara is a species of excavates. It has a PAS-positive surface layer and forms cysts in culture.
Dinophysis acuminata is a marine plankton species of dinoflagellates that is found in coastal waters of the north Atlantic and Pacific oceans. The genus Dinophysis includes both phototrophic and heterotrophic species. D. acuminata is one of several phototrophic species of Dinophysis classed as toxic, as they produce okadaic acid which can cause diarrhetic shellfish poisoning (DSP). Okadiac acid is taken up by shellfish and has been found in the soft tissue of mussels and the liver of flounder species. When contaminated animals are consumed, they cause severe diarrhoea. D. acuminata blooms are constant threat to and indication of diarrhoeatic shellfish poisoning outbreaks.

Mesodinium rubrum is a species of ciliates. It constitutes a plankton community and is found throughout the year, most abundantly in spring and fall, in coastal areas. Although discovered in 1908, its scientific importance came into light in the late 1960s when it attracted scientists by the recurrent red colouration it caused by forming massive blooms, that cause red tides in the oceans.
Karyoklepty is a strategy for cellular evolution, whereby a predator cell appropriates the nucleus of a cell from another organism to supplement its own biochemical capabilities.

Dinoflagellates are eukaryotic plankton, existing in marine and freshwater environments. Previously, dinoflagellates had been grouped into two categories, phagotrophs and phototrophs. Mixotrophs, however include a combination of phagotrophy and phototrophy. Mixotrophic dinoflagellates are a sub-type of planktonic dinoflagellates and are part of the phylum Dinoflagellata. They are flagellated eukaryotes that combine photoautotrophy when light is available, and heterotrophy via phagocytosis. Dinoflagellates are one of the most diverse and numerous species of phytoplankton, second to diatoms.

Licnophora is a genus of ciliates in the family Licnophoridae. They typically have an hourglass-like shape and live as ectocommensals on marine animals.
Arve Elvebakk is a Norwegian mycologist and professor working from the Arctic University of Norway in Tromsø. He has published widely on Arctic biology, and climatology. Additionally, he collaborates with many mycologists across the world, and has published names for lichens in Australia, New Zealand, the South Pacific, and South America, and the Antarctic.
