The centromere links a pair of sister chromatids together during cell division. This constricted region of chromosome connects the sister chromatids, creating a short arm (p) and a long arm (q) on the chromatids. During mitosis, spindle fibers attach to the centromere via the kinetochore.

A couple of homologous chromosomes, or homologs, are a set of one maternal and one paternal chromosome that pair up with each other inside a cell during fertilization. Homologs have the same genes in the same loci, where they provide points along each chromosome that enable a pair of chromosomes to align correctly with each other before separating during meiosis. This is the basis for Mendelian inheritance, which characterizes inheritance patterns of genetic material from an organism to its offspring parent developmental cell at the given time and area.

A kinetochore is a disc-shaped protein structure associated with duplicated chromatids in eukaryotic cells where the spindle fibers attach during cell division to pull sister chromatids apart. The kinetochore assembles on the centromere and links the chromosome to microtubule polymers from the mitotic spindle during mitosis and meiosis. The term kinetochore was first used in a footnote in a 1934 Cytology book by Lester W. Sharp and commonly accepted in 1936. Sharp's footnote reads: "The convenient term kinetochore has been suggested to the author by J. A. Moore", likely referring to John Alexander Moore who had joined Columbia University as a freshman in 1932.

Separase, also known as separin, is a cysteine protease responsible for triggering anaphase by hydrolysing cohesin, which is the protein responsible for binding sister chromatids during the early stage of anaphase. In humans, separin is encoded by the ESPL1 gene.
Adenomatous polyposis coli (APC) also known as deleted in polyposis 2.5 (DP2.5) is a protein that in humans is encoded by the APC gene. The APC protein is a negative regulator that controls beta-catenin concentrations and interacts with E-cadherin, which are involved in cell adhesion. Mutations in the APC gene may result in colorectal cancer and desmoid tumors.
SMC complexes represent a large family of ATPases that participate in many aspects of higher-order chromosome organization and dynamics. SMC stands for Structural Maintenance of Chromosomes.

Cohesin is a protein complex that mediates sister chromatid cohesion, homologous recombination, and DNA looping. Cohesin is formed of SMC3, SMC1, SCC1 and SCC3. Cohesin holds sister chromatids together after DNA replication until anaphase when removal of cohesin leads to separation of sister chromatids. The complex forms a ring-like structure and it is believed that sister chromatids are held together by entrapment inside the cohesin ring. Cohesin is a member of the SMC family of protein complexes which includes Condensin, MukBEF and SMC-ScpAB.
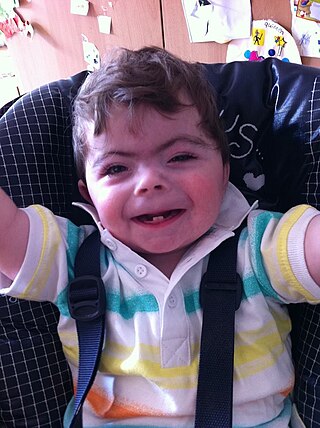
Cornelia de Lange syndrome (CdLS) is a genetic disorder. People with Cornelia de Lange syndrome experience a range of physical, cognitive, and medical challenges ranging from mild to severe. Cornelia de Lange syndrome has a widely varied phenotype, meaning people with the syndrome have varied features and challenges. The typical features of CdLS include thick or long eyebrows, a small nose, small stature, developmental delay, long or smooth philtrum, thin upper lip and downturned mouth.

Double-strand-break repair protein rad21 homolog is a protein that in humans is encoded by the RAD21 gene. RAD21, an essential gene, encodes a DNA double-strand break (DSB) repair protein that is evolutionarily conserved in all eukaryotes from budding yeast to humans. RAD21 protein is a structural component of the highly conserved cohesin complex consisting of RAD21, SMC1A, SMC3, and SCC3 [ STAG1 (SA1) and STAG2 (SA2) in multicellular organisms] proteins, involved in sister chromatid cohesion.

Structural maintenance of chromosomes protein 3 (SMC3) is a protein that in humans is encoded by the SMC3 gene. SMC3 is a subunit of the Cohesin complex which mediates sister chromatid cohesion, homologous recombination and DNA looping. Cohesin is formed of SMC3, SMC1, RAD21 and either SA1 or SA2. In humans, SMC3 is present in all cohesin complexes whereas there are multiple paralogs for the other subunits.

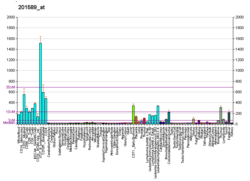
Cohesin subunit SA-2 (SA2) is a protein that in humans is encoded by the STAG2 gene. SA2 is a subunit of the Cohesin complex which mediates sister chromatid cohesion, homologous recombination and DNA looping. In somatic cells cohesin is formed of SMC3, SMC1, RAD21 and either SA1 or SA2 whereas in meiosis, cohesin is formed of SMC3, SMC1B, REC8 and SA3.

Wings apart-like protein homolog (WAPL) is a protein that in humans is encoded by the WAPAL gene. WAPL is a key regulator of the Cohesin complex which mediates sister chromatid cohesion, homologous recombination and DNA looping. Cohesin is formed of SMC3, SMC1, RAD21 and either SA1 or SA2. Cohesin has a ring-like arrangement and it is thought that it associates with the chromosome by entrapping it whether as a loop of DNA, a single strand or a pair of sister chromosomes. WAPL forms a complex with PDS5A or PDS5B and releases cohesin from DNA by opening the interface between SMC3 and RAD21.

Structural maintenance of chromosomes protein 4 (SMC-4) also known as chromosome-associated polypeptide C (CAP-C) or XCAP-C homolog is a protein that in humans is encoded by the SMC4 gene. SMC-4 is a core subunit of condensin I and II, large protein complexes involved in high order chromosome organization, including condensation and segregation. SMC-4 protein is commonly associated with the SMC-2 protein, another protein complex within the SMC protein family. SMC-4 dimerizes with SMC-2, creating the flexible and dynamic structure of the condensin holocomplex. An over-expression of the SMC-4 protein is shown to impact carcinogenesis.

Sister chromatid cohesion protein PDS5 homolog B(PDS5B) is a protein that in humans is encoded by the PDS5B gene. It is a regulatory subunit of the Cohesin complex which mediates sister chromatid cohesion, homologous recombination and DNA looping. The core cohesin complex is formed of SMC3, SMC1, RAD21 and either SA1 or SA2. PDS5 associates with WAPL to stimulate the release of cohesin from DNA but during DNA replication PDS5 promotes acetylation of SMC3 by ESCO1 and ESCO2.

Structural maintenance of chromosomes protein 5 is a protein encoded by the SMC5 gene in human.

Meiotic recombination protein REC8 homolog is a protein that in humans is encoded by the REC8 gene.
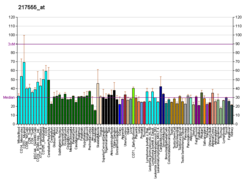
Cohesin subunit SA-1 (SA1) is a protein that in humans is encoded by the STAG1 gene. SA1 is a subunit of the Cohesin complex which mediates sister chromatid cohesion, homologous recombination and DNA looping. In somatic cells cohesin is formed of SMC3, SMC1, RAD21 and either SA1 or SA2 whereas in meiosis, cohesin is formed of SMC3, SMC1B, REC8 and SA3. There is a nonprofit community formed for those with a STAG1 Gene mutation at www.stag1gene.org.
Sister chromatid cohesion refers to the process by which sister chromatids are paired and held together during certain phases of the cell cycle. Establishment of sister chromatid cohesion is the process by which chromatin-associated cohesin protein becomes competent to physically bind together the sister chromatids. In general, cohesion is established during S phase as DNA is replicated, and is lost when chromosomes segregate during mitosis and meiosis. Some studies have suggested that cohesion aids in aligning the kinetochores during mitosis by forcing the kinetochores to face opposite cell poles.

Structural maintenance of chromosomes protein 1B (SMC-1B) is a protein that in humans is encoded by the SMC1B gene. SMC proteins engage in chromosome organization and can be broken into 3 groups based on function which are cohesins, condensins, and DNA repair. SMC-1B belongs to a family of proteins required for chromatid cohesion and DNA recombination during meiosis and mitosis. SMC1B protein appears to participate with other cohesins REC8, STAG3 and SMC3 in sister-chromatid cohesion throughout the whole meiotic process in human oocytes.

A topologically associating domain (TAD) is a self-interacting genomic region, meaning that DNA sequences within a TAD physically interact with each other more frequently than with sequences outside the TAD. The median size of a TAD in mouse cells is 880 kb, and they have similar sizes in non-mammalian species. Boundaries at both side of these domains are conserved between different mammalian cell types and even across species and are highly enriched with CCCTC-binding factor (CTCF) and cohesin. In addition, some types of genes appear near TAD boundaries more often than would be expected by chance.