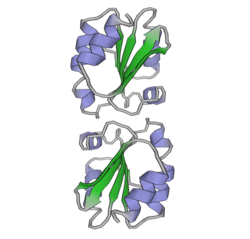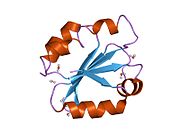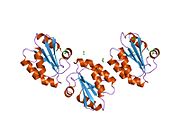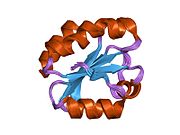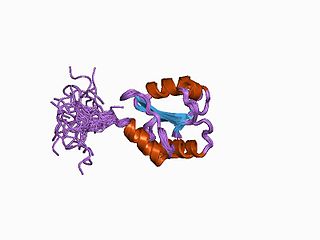
Protein disulfide isomerase, or PDI, is an enzyme in the endoplasmic reticulum (ER) in eukaryotes and the periplasm of bacteria that catalyzes the formation and breakage of disulfide bonds between cysteine residues within proteins as they fold. This allows proteins to quickly find the correct arrangement of disulfide bonds in their fully folded state, and therefore the enzyme acts to catalyze protein folding.
Thioredoxin reductases are enzymes that reduce thioredoxin (Trx). Two classes of thioredoxin reductase have been identified: one class in bacteria and some eukaryotes and one in animals. In bacteria TrxR also catalyzes the reduction of glutaredoxin like proteins known as NrdH. Both classes are flavoproteins which function as homodimers. Each monomer contains a FAD prosthetic group, a NADPH binding domain, and an active site containing a redox-active disulfide bond.
Ferredoxins are iron–sulfur proteins that mediate electron transfer in a range of metabolic reactions. The term "ferredoxin" was coined by D.C. Wharton of the DuPont Co. and applied to the "iron protein" first purified in 1962 by Mortenson, Valentine, and Carnahan from the anaerobic bacterium Clostridium pasteurianum.

Glutathione reductase (GR) also known as glutathione-disulfide reductase (GSR) is an enzyme that in humans is encoded by the GSR gene. Glutathione reductase catalyzes the reduction of glutathione disulfide (GSSG) to the sulfhydryl form glutathione (GSH), which is a critical molecule in resisting oxidative stress and maintaining the reducing environment of the cell. Glutathione reductase functions as dimeric disulfide oxidoreductase and utilizes an FAD prosthetic group and NADPH to reduce one molar equivalent of GSSG to two molar equivalents of GSH:

Vitamin K epoxide reductase (VKOR) is an enzyme that reduces vitamin K after it has been oxidised in the carboxylation of glutamic acid residues in blood coagulation enzymes. VKOR is a member of a large family of predicted enzymes that are present in vertebrates, Drosophila, plants, bacteria and archaea. In some plant and bacterial homologues, the VKOR domain is fused with domains of the thioredoxin family of oxidoreductases.

Glutaredoxins are small redox enzymes of approximately one hundred amino-acid residues that use glutathione as a cofactor. In humans this oxidation repair enzyme is also known to participate in many cellular functions, including redox signaling and regulation of glucose metabolism. Glutaredoxins are oxidized by substrates, and reduced non-enzymatically by glutathione. In contrast to thioredoxins, which are reduced by thioredoxin reductase, no oxidoreductase exists that specifically reduces glutaredoxins. Instead, glutaredoxins are reduced by the oxidation of glutathione. Reduced glutathione is then regenerated by glutathione reductase. Together these components compose the glutathione system.
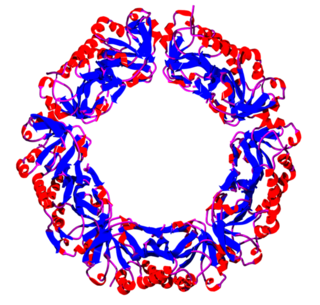
Peroxiredoxins are a ubiquitous family of antioxidant enzymes that also control cytokine-induced peroxide levels and thereby mediate signal transduction in mammalian cells. The family members in humans are PRDX1, PRDX2, PRDX3, PRDX4, PRDX5, and PRDX6. The physiological importance of peroxiredoxins is indicated by their relative abundance. Their function is the reduction of peroxides, specifically hydrogen peroxide, alkyl hydroperoxides, and peroxynitrite.

Apoptosis signal-regulating kinase 1 (ASK1) also known as mitogen-activated protein kinase 5 (MAP3K5) is a member of MAP kinase family and as such a part of mitogen-activated protein kinase pathway. It activates c-Jun N-terminal kinase (JNK) and p38 mitogen-activated protein kinases in a Raf-independent fashion in response to an array of stresses such as oxidative stress, endoplasmic reticulum stress and calcium influx. ASK1 has been found to be involved in cancer, diabetes, rheumatoid arthritis, cardiovascular and neurodegenerative diseases.
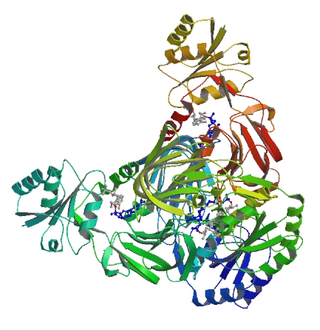
NAD+ kinase (EC 2.7.1.23, NADK) is an enzyme that converts nicotinamide adenine dinucleotide (NAD+) into NADP+ through phosphorylating the NAD+ coenzyme. NADP+ is an essential coenzyme that is reduced to NADPH primarily by the pentose phosphate pathway to provide reducing power in biosynthetic processes such as fatty acid biosynthesis and nucleotide synthesis. The structure of the NADK from the archaean Archaeoglobus fulgidus has been determined.
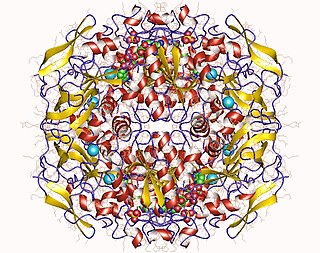
In enzymology, a formaldehyde dehydrogenase (EC 1.2.1.46) is an enzyme that catalyzes the chemical reaction

Protein disulfide-isomerase, also known as the beta-subunit of prolyl 4-hydroxylase (P4HB), is an enzyme that in humans encoded by the P4HB gene. The human P4HB gene is localized in chromosome 17q25. Unlike other prolyl 4-hydroxylase family proteins, this protein is multifunctional and acts as an oxidoreductase for disulfide formation, breakage, and isomerization. The activity of P4HB is tightly regulated. Both dimer dissociation and substrate binding are likely to enhance its enzymatic activity during the catalysis process.

Thioredoxin-interacting protein is a protein that in humans is encoded by the TXNIP gene.

Thioredoxin-dependent peroxide reductase, mitochondrial is an enzyme that in humans is encoded by the PRDX3 gene. It is a member of the peroxiredoxin family of antioxidant enzymes.

Glutaredoxin 2 (GLRX2) is an enzyme that in humans encoded by the GLRX2 gene. GLRX2, also known as GRX2, is a glutaredoxin family protein and a thiol-disulfide oxidoreductase that maintains cellular thiol homeostasis. This gene consists of four exons and three introns, spanned 10 kilobase pairs, and localized to chromosome 1q31.2–31.3.
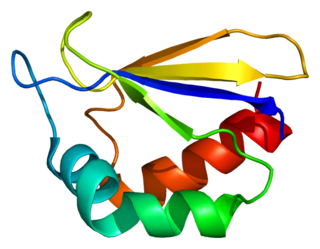
NADH dehydrogenase [ubiquinone] 1 alpha subcomplex subunit 2 is a protein that in humans is encoded by the NDUFA2 gene. The NDUFA2 protein is a subunit of NADH dehydrogenase (ubiquinone), which is located in the mitochondrial inner membrane and is the largest of the five complexes of the electron transport chain. Mutations in the NDUFA2 gene are associated with Leigh's syndrome.
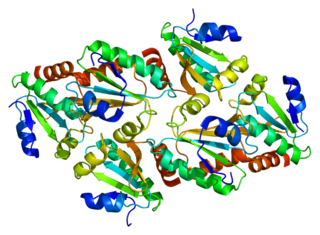
Thioredoxin, mitochondrial also known as thioredoxin-2 is a protein that in humans is encoded by the TXN2 gene on chromosome 22. This nuclear gene encodes a mitochondrial member of the thioredoxin family, a group of small multifunctional redox-active proteins. The encoded protein may play important roles in the regulation of the mitochondrial membrane potential and in protection against oxidant-induced apoptosis.
In biochemistry, S-nitrosylation is the covalent attachment of a nitric oxide group to a cysteine thiol within a protein to form an S-nitrosothiol (SNO). S-Nitrosylation has diverse regulatory roles in bacteria, yeast and plants and in all mammalian cells. It thus operates as a fundamental mechanism for cellular signaling across phylogeny and accounts for the large part of NO bioactivity.

Ferredoxin-thioredoxin reductase EC 1.8.7.2, systematic name ferredoxin:thioredoxin disulfide oxidoreductase, is a [4Fe-4S] protein that plays an important role in the ferredoxin/thioredoxin regulatory chain. It catalyzes the following reaction:

Methionine sulfoxide is the organic compound with the formula CH3S(O)CH2CH2CH(NH2)CO2H. It is an amino acid that occurs naturally although it is formed post-translationally.

Protein disulfide isomerase family A member 2 is a protein that in humans is encoded by the PDIA2 gene.