Turtles, or testudines, are reptiles of the order Testudines, characterized by a special shell developed mainly from their ribs. Modern turtles are divided into two major groups, the Pleurodira and Cryptodira, which differ in the way the head retracts. There are 360 living and recently extinct species of turtles, including land-dwelling tortoises and freshwater terrapins. They are found on most continents, some islands and, in the case of sea turtles, much of the ocean. Like other amniotes they breathe air and do not lay eggs underwater, although many species live in or around water.

Chelidae is one of three living families of the turtle suborder Pleurodira, and are commonly called Austro-South American side-neck turtles. The family is distributed in Australia, New Guinea, parts of Indonesia, and throughout most of South America. It is a large family of turtles with a significant fossil history dating back to the Cretaceous. The family is entirely Gondwanan in origin, with no members found outside Gondwana, either in the present day or as a fossil.

Proganochelys is a genus of extinct, primitive stem-turtle. Proganochelys was named by Georg Baur in 1887 as the oldest turtle in existence at the time. The name Proganochelys comes from the Greek word ganos meaning 'brightness', combined with prefix pro, 'before', and Greek base chelys meaning 'turtle'. Proganochelys is believed to have been around 1 meter in size and herbivorous in nature. Proganochelys was known as the most primitive stem-turtle for over a century, until the novel discovery of Odontochelys in 2008. Odontochelys and Proganochelys share unique primitive features that are not found in Casichelydia, such as teeth on the pterygoid and vomer and a plate-like coracoid.

The Amboina box turtle or Southeast Asian box turtle is a species of Asian box turtle widely distributed across Southeast Asia. It is native to the Asian mainland from northeast India, through Bangladesh, Burma and Thailand, across Laos, Cambodia, Vietnam, and Malaysia. It is also found on the archipelagos of Indonesia and the Philippines.

The Cochin forest cane turtle, also known as Kavalai forest turtle, forest cane turtle or simply cane turtle, is a rare turtle from the Western Ghats of India. Described in 1912, its type locality is given as "Near Kavalai in the Cochin State Forests, inhabiting dense forest, at an elevation of about 1500 feet above sea level". Only two specimens were found at that time, and no scientist saw this turtle for the next 70 years. It was rediscovered in 1982, and since then a number of specimens have been found and some studies have been conducted about its phylogeny and ecology.

The Indian roofed turtle is a species of turtle in the family Geoemydidae. It can be distinguished by the distinct "roof" at the topmost part of the shell. It is found in the major rivers of South Asia. It is a common pet in the Indian Subcontinent.

Stupendemys is an extinct genus of freshwater side-necked turtle, belonging to the family Podocnemididae. It is the largest freshwater turtle known to have existed, with a carapace over 2 meters long. Its fossils have been found in northern South America, in rocks dating from the Middle Miocene to the very start of the Pliocene, about 13 to 5 million years ago. Male specimens are known to have possessed bony horns growing from the front edges of the shell and the discovery of the fossil of a young adult shows that the carapace of these turtles flattens with age. A fossil skull described in 2021 indicates that Stupendemys was a generalist feeder.

The Pleurodira are one of the two living suborders of turtles, the other being the Cryptodira. The division between these two suborders represents a very deep evolutionary divide between two very different types of turtles. The physical differences between them, although anatomical and largely internal, are nonetheless significant, and the zoogeographic implications of them are substantial. The Pleurodira are known more commonly as the side-necked turtles and the name Pleurodira quite literally translates to side neck, whereas the Cryptodira are known as hidden-necked turtles. The Pleurodira turtles are currently restricted to freshwater habitats in the Southern Hemisphere, largely to Australia, South America, and Africa. Within the Pleurodira, three living families are represented: Chelidae, also known as the Austro-South American side-necked turtles, the Pelomedusidae, also known as the African mud terrapins, and the Podocnemididae, also known as the American side-neck river turtles. However, they were cosmopolitan clade during the Cretaceous and most of the Cenozoic, and even occurred in marine environments around the world.

Pelusios is a genus of African side-necked turtles. With 17 described species, it is one of the most diverse genera of the turtle order (Testudines).

The big-headed pantanal swamp turtle or pantanal swamp turtle is a species of turtle in the family Chelidae found in Brazil, Bolivia, Argentina, and Paraguay.

The yellow-footed tortoise, also known as the Brazilian giant tortoise, is a species of tortoise in the family Testudinidae and is closely related to the red-footed tortoise. It is found in the Amazon Basin of South America. The species name has often been misspelled as denticulata, an error introduced in the 1980s when Chelonoidis was elevated to genus and mistakenly treated as feminine, an error recognized and fixed in 2017.
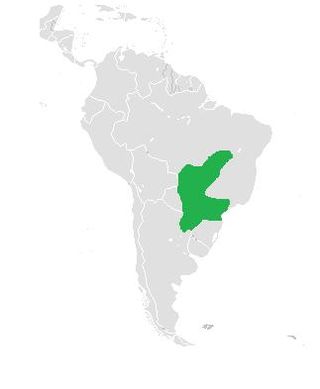
Vanderhaege's toad-headed turtle is a species of turtle in the family Chelidae. The species is endemic to South America.

Odontochelys semitestacea is a Late Triassic relative of turtles before Pappochelys was discovered and Eunotosaurus was redescribed, Odontochelys was considered the oldest undisputed member of Pantestudines. It is the only known species in the genus Odontochelys and the family Odontochelyidae.

Eunotosaurus is an extinct genus of amniote, possibly a close relative of turtles. Eunotosaurus lived in the late Middle Permian and fossils can be found in the Karoo Supergroup of South Africa. Eunotosaurus resided in the swamps of what is now southern Africa. Its ribs were wide and flat, forming broad plates similar to a primitive turtle shell, and the vertebrae were nearly identical to those of some turtles. Accordingly, it is often considered as a possible transitional fossil between turtles and their prehistoric ancestors. However, it is possible that these turtle-like features evolved independently of the same features in turtles, since other anatomical studies and phylogenetic analyses suggest that Eunotosaurus may instead have been a parareptile, an early-diverging neodiapsid unrelated to turtles, or a synapsid.

Cyclemys is a genus of freshwater turtles, commonly referred to as Asian leaf turtles, from the family Geoemydidae. The genus occurs throughout Southeast and South Asia, and currently contains seven species.

Cyamodontoidea is an extinct superfamily of placodont marine reptiles from the Triassic period. It is one of the two main groups of placodonts, the other being Placodontoidea. Cyamodontoids are distinguished from placodontoids by their large shells, formed from fused bony plates called osteoderms and superficially resembling the shells of turtles. Cyamodontoids also have distinctive skulls with narrow, often toothless jaws and wide, flaring temporal regions behind the eyes. Two large temporal openings are positioned at the top of the back of the skull, an arrangement that is known as the euryapsid condition and seen throughout Sauropterygia, the marine reptile group to which placodonts belong. Cyamodontoids are also distinguished by their large crushing teeth, which grow from the palatine bones on the roof of the mouth.
Chinlechelys is an extinct genus of stem-turtle belonging to Testudinata. It lived in the Norian age of the Late Triassic and is the oldest turtle known from North America. Among turtles it is unique, mostly because of its very thin shell. The type and only species, C. tenertesta, was named and described with the genus by Walter G. Joyce et al. in 2009. It was probably terrestrial, and was found by Joyce et al. to be closely related to Proganochelys, another terrestrial testudinatan.

Pantestudines or Pan-Testudines is the proposed group of all reptiles more closely related to turtles than to any other living animal. It includes both modern turtles and all of their extinct relatives. Pantestudines with a complete shell are placed in the clade Testudinata.

Pappochelys is an extinct genus of diapsid reptile possibly related to turtles. The genus contains only one species, Pappochelys rosinae, from the Middle Triassic of Germany, which was named by paleontologists Rainer Schoch and Hans-Dieter Sues in 2015. The discovery of Pappochelys provides strong support for the placement of turtles within Diapsida, a hypothesis that has long been suggested by molecular data, but never previously by the fossil record. It is morphologically intermediate between the definite stem-turtle Odontochelys from the Late Triassic of China and Eunotosaurus, a reptile from the Middle Permian of South Africa.
Eorhynchochelys is an extinct genus of stem-turtle from the Late Triassic Xiaowa Formation of southwestern China.