
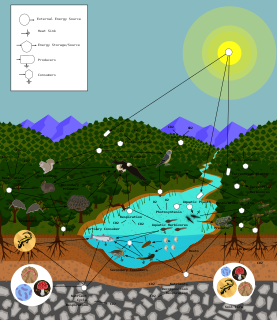
Ecology is the study of the relationships between living organisms, including humans, and their physical environment. Ecology considers organisms at the individual, population, community, ecosystem, and biosphere level. Ecology overlaps with the closely related sciences of biogeography, evolutionary biology, genetics, ethology, and natural history. Ecology is a branch of biology, and it is not synonymous with environmentalism.
Theoretical ecology is the scientific discipline devoted to the study of ecological systems using theoretical methods such as simple conceptual models, mathematical models, computational simulations, and advanced data analysis. Effective models improve understanding of the natural world by revealing how the dynamics of species populations are often based on fundamental biological conditions and processes. Further, the field aims to unify a diverse range of empirical observations by assuming that common, mechanistic processes generate observable phenomena across species and ecological environments. Based on biologically realistic assumptions, theoretical ecologists are able to uncover novel, non-intuitive insights about natural processes. Theoretical results are often verified by empirical and observational studies, revealing the power of theoretical methods in both predicting and understanding the noisy, diverse biological world.

In ecology, a niche is the match of a species to a specific environmental condition. It describes how an organism or population responds to the distribution of resources and competitors and how it in turn alters those same factors. "The type and number of variables comprising the dimensions of an environmental niche vary from one species to another [and] the relative importance of particular environmental variables for a species may vary according to the geographic and biotic contexts".
This glossary of ecology is a list of definitions of terms and concepts in ecology and related fields. For more specific definitions from other glossaries related to ecology, see Glossary of biology, Glossary of evolutionary biology, and Glossary of environmental science.

In ecology, the competitive exclusion principle, sometimes referred to as Gause's law, is a proposition that two species which compete for the same limited resource cannot coexist at constant population values. When one species has even the slightest advantage over another, the one with the advantage will dominate in the long term. This leads either to the extinction of the weaker competitor or to an evolutionary or behavioral shift toward a different ecological niche. The principle has been paraphrased in the maxim "complete competitors can not coexist".

In biology, polymorphism is the occurrence of two or more clearly different morphs or forms, also referred to as alternative phenotypes, in the population of a species. To be classified as such, morphs must occupy the same habitat at the same time and belong to a panmictic population.
Evolutionary ecology lies at the intersection of ecology and evolutionary biology. It approaches the study of ecology in a way that explicitly considers the evolutionary histories of species and the interactions between them. Conversely, it can be seen as an approach to the study of evolution that incorporates an understanding of the interactions between the species under consideration. The main subfields of evolutionary ecology are life history evolution, sociobiology, the evolution of interspecific interactions and the evolution of biodiversity and of ecological communities.

Charles Sutherland Elton was an English zoologist and animal ecologist. He is associated with the development of population and community ecology, including studies of invasive organisms.
In ecology, niche differentiation refers to the process by which competing species use the environment differently in a way that helps them to coexist. The competitive exclusion principle states that if two species with identical niches compete, then one will inevitably drive the other to extinction. This rule also states that two species cannot occupy the same exact niche in a habitat and coexist together, at least in a stable manner. When two species differentiate their niches, they tend to compete less strongly, and are thus more likely to coexist. Species can differentiate their niches in many ways, such as by consuming different foods, or using different areas of the environment.
The hypothesis of effective evolutionary time attempts to explain gradients, in particular latitudinal gradients, in species diversity. It was originally named "time hypothesis".

Species use restricted ecological niches, and the niches of all species are segregated, often with much overlap, by the use of different habitats, different geographic areas and seasons, and different food resources, to mention only a few of the many niche dimensions. The causes of niche restriction and segregation are important problems in evolutionary ecology.
The following outline is provided as an overview of and topical guide to ecology:

In ecology, a community is a group or association of populations of two or more different species occupying the same geographical area at the same time, also known as a biocoenosis, biotic community, biological community, ecological community, or life assemblage. The term community has a variety of uses. In its simplest form it refers to groups of organisms in a specific place or time, for example, "the fish community of Lake Ontario before industrialization".
Source–sink dynamics is a theoretical model used by ecologists to describe how variation in habitat quality may affect the population growth or decline of organisms.
Ecological forecasting uses knowledge of physics, ecology and physiology to predict how ecological populations, communities, or ecosystems will change in the future in response to environmental factors such as climate change. The goal of the approach is to provide natural resource managers with information to anticipate and respond to short and long-term climate conditions.

Species distribution modelling (SDM), also known as environmental(or ecological) niche modelling (ENM), habitat modelling, predictive habitat distribution modelling, and range mapping uses computer algorithms to predict the distribution of a species across geographic space and time using environmental data. The environmental data are most often climate data, but can include other variables such as soil type, water depth, and land cover. SDMs are used in several research areas in conservation biology, ecology and evolution. These models can be used to understand how environmental conditions influence the occurrence or abundance of a species, and for predictive purposes. Predictions from an SDM may be of a species’ future distribution under climate change, a species’ past distribution in order to assess evolutionary relationships, or the potential future distribution of an invasive species. Predictions of current and/or future habitat suitability can be useful for management applications.

Klaus Rohde is a German biologist at the University of New England (UNE), Australia, known particularly for his work on marine parasitology, evolutionary ecology/zoogeography, and phylogeny/ultrastructure of lower invertebrates.

Ecological fitting is "the process whereby organisms colonize and persist in novel environments, use novel resources or form novel associations with other species as a result of the suites of traits that they carry at the time they encounter the novel condition". It can be understood as a situation in which a species' interactions with its biotic and abiotic environment seem to indicate a history of coevolution, when in actuality the relevant traits evolved in response to a different set of biotic and abiotic conditions.
The term phylogenetic niche conservatism has seen increasing use in recent years in the scientific literature, though the exact definition has been a matter of some contention. Fundamentally, phylogenetic niche conservatism refers to the tendency of species to retain their ancestral traits. When defined as such, phylogenetic niche conservatism is therefore nearly synonymous with phylogenetic signal. The point of contention is whether or not "conservatism" refers simply to the tendency of species to resemble their ancestors, or implies that "closely related species are more similar than expected based on phylogenetic relationships". If the latter interpretation is employed, then phylogenetic niche conservatism can be seen as an extreme case of phylogenetic signal, and implies that the processes which prevent divergence are in operation in the lineage under consideration. Despite efforts by Jonathan Losos to end this habit, however, the former interpretation appears to frequently motivate scientific research. In this case, phylogenetic niche conservatism might best be considered a form of phylogenetic signal reserved for traits with broad-scale ecological ramifications. Thus, phylogenetic niche conservatism is usually invoked with regards to closely related species occurring in similar environments.
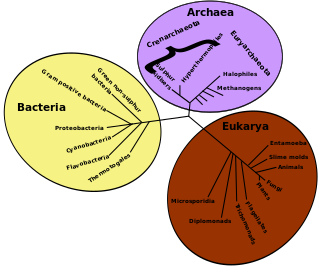
This glossary of evolutionary biology is a list of definitions of terms and concepts used in the study of evolutionary biology, population biology, speciation, and phylogenetics, as well as sub-disciplines and related fields. For additional terms from related glossaries, see Glossary of genetics, Glossary of ecology, and Glossary of biology.

