
Asexual reproduction is a type of reproduction that does not involve the fusion of gametes or change in the number of chromosomes. The offspring that arise by asexual reproduction from either unicellular or multicellular organisms inherit the full set of genes of their single parent and thus the newly created individual is genetically and physically similar to the parent or an exact clone of the parent. Asexual reproduction is the primary form of reproduction for single-celled organisms such as archaea and bacteria. Many eukaryotic organisms including plants, animals, and fungi can also reproduce asexually. In vertebrates, the most common form of asexual reproduction is parthenogenesis, which is typically used as an alternative to sexual reproduction in times when reproductive opportunities are limited. Komodo dragons and some monitor lizards can reproduce asexually.

Meiosis is a special type of cell division of germ cells and apicomplexans in sexually-reproducing organisms that produces the gametes, the sperm or egg cells. It involves two rounds of division that ultimately result in four cells, each with only one copy of each chromosome (haploid). Additionally, prior to the division, genetic material from the paternal and maternal copies of each chromosome is crossed over, creating new combinations of code on each chromosome. Later on, during fertilisation, the haploid cells produced by meiosis from a male and a female will fuse to create a zygote, a cell with two copies of each chromosome again.

Volvox is a polyphyletic genus of chlorophyte green algae in the family Volvocaceae. Volvox species form spherical colonies of up to 50,000 cells, and for this reason they are sometimes called globe algae. They live in a variety of freshwater habitats, and were first reported by Antonie van Leeuwenhoek in 1700. Volvox diverged from unicellular ancestors approximately 200 million years ago.
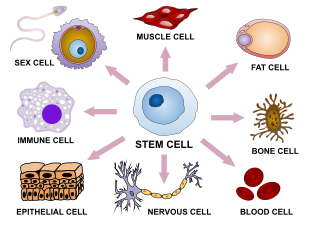
Cellular differentiation is the process in which a stem cell changes from one type to a differentiated one. Usually, the cell changes to a more specialized type. Differentiation happens multiple times during the development of a multicellular organism as it changes from a simple zygote to a complex system of tissues and cell types. Differentiation continues in adulthood as adult stem cells divide and create fully differentiated daughter cells during tissue repair and during normal cell turnover. Some differentiation occurs in response to antigen exposure. Differentiation dramatically changes a cell's size, shape, membrane potential, metabolic activity, and responsiveness to signals. These changes are largely due to highly controlled modifications in gene expression and are the study of epigenetics. With a few exceptions, cellular differentiation almost never involves a change in the DNA sequence itself. However, metabolic composition does get altered quite dramatically where stem cells are characterized by abundant metabolites with highly unsaturated structures whose levels decrease upon differentiation. Thus, different cells can have very different physical characteristics despite having the same genome.
A cell type is a classification used to identify cells that share morphological or phenotypical features. A multicellular organism may contain cells of a number of widely differing and specialized cell types, such as muscle cells and skin cells, that differ both in appearance and function yet have identical genomic sequences. Cells may have the same genotype, but belong to different cell types due to the differential regulation of the genes they contain. Classification of a specific cell type is often done through the use of microscopy. Recent developments in single cell RNA sequencing facilitated classification of cell types based on shared gene expression patterns. This has led to the discovery of many new cell types in e.g. mouse cortex, hippocampus, dorsal root ganglion and spinal cord.
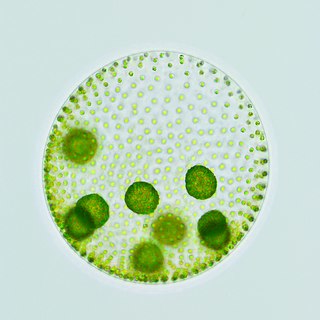
The Volvocaceae are a family of unicellular or colonial biflagellates, including the typical genus Volvox. The family was named by Ehrenberg in 1834, and is known in older classifications as the Volvocidae. All species are colonial and inhabit freshwater environments.
In cellular biology, a somatic cell, or vegetal cell, is any biological cell forming the body of a multicellular organism other than a gamete, germ cell, gametocyte or undifferentiated stem cell. Somatic cells compose the body of an organism and divide through mitosis.

A unicellular organism, also known as a single-celled organism, is an organism that consists of a single cell, unlike a multicellular organism that consists of multiple cells. Organisms fall into two general categories: prokaryotic organisms and eukaryotic organisms. Most prokaryotes are unicellular and are classified into bacteria and archaea. Many eukaryotes are multicellular, but some are unicellular such as protozoa, unicellular algae, and unicellular fungi. Unicellular organisms are thought to be the oldest form of life, with early protocells possibly emerging 3.8–4.8 billion years ago.
A multicellular organism is an organism that consists of more than one cell, in contrast to unicellular organism. All species of animals, land plants and most fungi are multicellular, as are many algae, whereas a few organisms are partially uni- and partially multicellular, like slime molds and social amoebae such as the genus Dictyostelium.

In biology, a biological life cycle is a series of stages of the life of an organism, that begins as a zygote, often in an egg, and concludes as an adult that reproduces, producing an offspring in the form of a new zygote which then itself goes through the same series of stages, the process repeating in a cyclic fashion.

The green algae are a group of chlorophyll-containing autotrophic eukaryotes consisting of the phylum Prasinodermophyta and its unnamed sister group that contains the Chlorophyta and Charophyta/Streptophyta. The land plants (Embryophytes) have emerged deep in the Charophyte alga as a sister of the Zygnematophyceae. Since the realization that the Embryophytes emerged within the green algae, some authors are starting to include them. The completed clade that includes both green algae and embryophytes is monophyletic and is referred to as the clade Viridiplantae and as the kingdom Plantae. The green algae include unicellular and colonial flagellates, most with two flagella per cell, as well as various colonial, coccoid and filamentous forms, and macroscopic, multicellular seaweeds. There are about 22,000 species of green algae, many of which live most of their lives as single cells, while other species form coenobia (colonies), long filaments, or highly differentiated macroscopic seaweeds.

In biology and genetics, the germline is the population of a multicellular organism's cells that develop into germ cells. In other words, they are the cells that form gametes, which can come together to form a zygote. They differentiate in the gonads from primordial germ cells into gametogonia, which develop into gametocytes, which develop into the final gametes. This process is known as gametogenesis.

Sexual reproduction is an adaptive feature which is common to almost all multicellular organisms and various unicellular organisms. Currently, the adaptive advantage of sexual reproduction is widely regarded as a major unsolved problem in biology. As discussed below, one prominent theory is that sex evolved as an efficient mechanism for producing variation, and this had the advantage of enabling organisms to adapt to changing environments. Another prominent theory, also discussed below, is that a primary advantage of outcrossing sex is the masking of the expression of deleterious mutations. Additional theories concerning the adaptive advantage of sex are also discussed below. Sex does, however, come with a cost. In reproducing asexually, no time nor energy needs to be expended in choosing a mate and, if the environment has not changed, then there may be little reason for variation, as the organism may already be well-adapted. However, very few environments have not changed over the millions of years that reproduction has existed. Hence it is easy to imagine that being able to adapt to changing environment imparts a benefit. Sex also halves the amount of offspring a given population is able to produce. Sex, however, has evolved as the most prolific means of species branching into the tree of life. Diversification into the phylogenetic tree happens much more rapidly via sexual reproduction than it does by way of asexual reproduction.
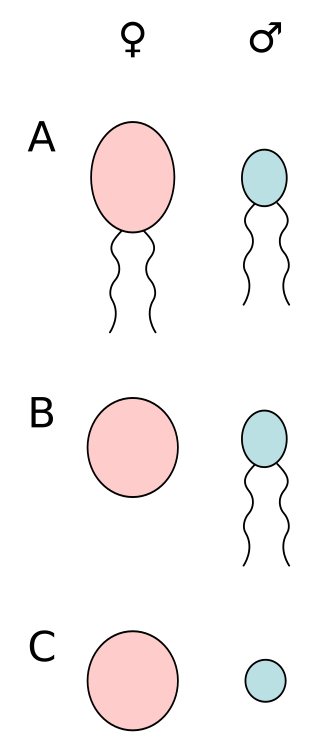
Anisogamy is a form of sexual reproduction that involves the union or fusion of two gametes that differ in size and/or form. The smaller gamete is male, a sperm cell, whereas the larger gamete is female, typically an egg cell. Anisogamy is predominant among multicellular organisms. In both plants and animals, gamete size difference is the fundamental difference between females and males.

In microbiology, genetics, cell biology, and molecular biology, competence is the ability of a cell to alter its genetics by taking up extracellular ("naked") DNA from its environment in the process called transformation. Competence may be differentiated between natural competence, a genetically specified ability of bacteria which is thought to occur under natural conditions as well as in the laboratory, and induced or artificial competence, which arises when cells in laboratory cultures are treated to make them transiently permeable to DNA. Competence allows for rapid adaptation and DNA repair of the cell. This article primarily deals with natural competence in bacteria, although information about artificial competence is also provided.

Gonium is a genus of colonial algae, a member of the order Chlamydomonadales. Typical colonies have 4 to 16 cells, all the same size, arranged in a flat plate, with no anterior-posterior differentiation. In a colony of 16 cells, four are in the center, and the other 12 are on the four sides, three each. A description by G.M. Smith :
Gonium Mueller 1773: Colonies of 4-8-16 cells arranged in a flat quadrangular plate and embedded in a common gelatinous matrix or connected by broad gelatinous strands. Cells ovoid to pyriform, with a single cup-shaped chloroplast containing one pyrenoid. Each cell with two cilia of equal length, contractile vacuoles at the base of the cilia, and an eyespot. Four- and eight-celled colonies with the cilia on the same side ; sixteen-celled colonies with the four central cells having their cilia on the same side and the twelve marginal cells with radially arranged cilia.
Asexual reproduction by simultaneous division of all cells in the colony to form autocolonies, or by a formation of 2-4 zoospores in each cell.
Sexual reproduction isogamous, by a fusion of biciliatezoogametes.

Sexual reproduction is a type of reproduction that involves a complex life cycle in which a gamete with a single set of chromosomes combines with another gamete to produce a zygote that develops into an organism composed of cells with two sets of chromosomes (diploid). This is typical in animals, though the number of chromosome sets and how that number changes in sexual reproduction varies, especially among plants, fungi, and other eukaryotes.

Ectocarpus siliculosus is a filamentous brown alga. Its genome was the first brown macroalgal genome to be sequenced, with the expectation that E. siliculosus will serve as a genetic and genomic model for brown macroalgae.
The origin and function of meiosis are currently not well understood scientifically, and would provide fundamental insight into the evolution of sexual reproduction in eukaryotes. There is no current consensus among biologists on the questions of how sex in eukaryotes arose in evolution, what basic function sexual reproduction serves, and why it is maintained, given the basic two-fold cost of sex. It is clear that it evolved over 1.2 billion years ago, and that almost all species which are descendants of the original sexually reproducing species are still sexual reproducers, including plants, fungi, and animals.
Germ-Soma Differentiation is the process by which organisms develop distinct germline and somatic cells. The development of cell differentiation has been one of the critical aspects of the evolution of multicellularity and sexual reproduction in organisms. Multicellularity has evolved upwards of 25 times, and due to this there is great possibility that multiple factors have shaped the differentiation of cells. There are three general types of cells: germ cells, somatic cells, and stem cells. Germ cells lead to the production of gametes, while somatic cells perform all other functions within the body. Within the broad category of somatic cells, there is further specialization as cells become specified to certain tissues and functions. In addition, stem cell are undifferentiated cells which can develop into a specialized cell and are the earliest type of cell in a cell lineage. Due to the differentiation in function, somatic cells are found ony in multicellular organisms, as in unicellular ones the purposes of somatic and germ cells are consolidated in one cell.
