CUG triplet repeat, RNA binding protein 1, also known as CUGBP1, is a protein which in humans is encoded by the CUGBP1 gene. [3] [4] [5]
CUG triplet repeat, RNA binding protein 1, also known as CUGBP1, is a protein which in humans is encoded by the CUGBP1 gene. [3] [4] [5]
Members of the CELF/BRUNOL protein family contain two N-terminal RNA recognition motif (RRM) domains, one C-terminal RRM domain, and a divergent segment of 160-230 aa between the second and third RRM domains. Members of this protein family regulate pre-mRNA alternative splicing and may also be involved in mRNA editing, and translation. This gene may play a role in myotonic dystrophy type 1 (DM1) via interactions with the dystrophia myotonica-protein kinase (DMPK) gene. Alternative splicing results in multiple transcript variants encoding different isoforms. [3]
It is estimated that 5 to 8% of human mRNAs are unstable because of mRNA instability elements in their 3' untranslated regions (3'UTR). [6] A number of such elements have been called AU-rich elements (AREs). It is now known that AREs are binding sites for RNA-binding proteins that target mRNAs to rapid degradation. However, only few of the proteins reported to bind AREs were demonstrated to play a role in mRNA degradation. A shared feature of these proteins is to bind only to a subclass of the known AREs that contain the pentamer AUUUA. A convergent effort of several research teams now adds CUGBP1 (CUG binding protein 1) to the short list of ARE-Binding proteins that control mRNA stability, with the peculiarity that it binds to non-AUUUA AREs. CUGBP1 has been involved both as a key regulator of human myotonic dystrophy 1 (DM1) and more recently as a regulator of human papilloma virus mRNA expression. [7]
Evidence for CUGBP1 acting as a RNA degradation factor came first from the Xenopus model. Xenopus CUGBP1 (xCUGBP1, formerly known as EDEN-BP) was identified in 1998 [8] for its ability to bind specifically to a GU-rich element (Embryonic deadenylation element EDEN) located in the 3'UTRs of some mRNAs that are rapidly deadenylated and translationally repressed after fertilization in early development. Because deadenylation is often the rate limiting step of mRNA degradation the enhancement of deadenylation increases mRNA turnover. [9]
Human CUGBP1 (hCUGBP1) had been previously identified by Timchenko and colleagues [4] for its ability to bind to CUG repeats located in the DMPK 3'UTR. A large amount of work has since described the role of hCUGBP1 on control of alternative splicing and will not be discussed here. [10] The demonstration that hCUGBP1 is involved in the control of mRNA deadenylation and instability like xCUGBP1 came next. In mammalian cell extract as well as in xenopus egg extracts, depletion and rescue experiments showed that specific binding of CUGBP1 to the 3'UTR of mRNA is required for the targeted specific deadenylation to occur. [11] In rescue experiments in xenopus egg extracts, the recombinant human protein can replace the xenopus one making them functional homolog. [12] Furthermore, the Poly(A) ribonuclease PARN was shown to interact with CUGBP1. [13] In human cells, tethering of hCUGBP1 to a mRNA decreases its steadystate suggesting the destabilization of the mRNA. [14] The first human mRNA reported to be targeted to rapid deadenylation and degradation by CUGBP1 is the oncogene c-jun. Years ago, it was shown that the class III ARE (devoid of any AUUUA motif) of the human c-jun oncogene directed rapid deadenylation and degradation to a reporter mRNA. [15] Both xCUGBP1 and hCUGBP1 were shown to specifically bind to c-jun ARE. [11] The binding of CUGBP1 to the 3'UTR of mRNAs bearing GU-rich element would target these mRNAs for rapid deadenylation by PARN and subsequent degradation. This was recently demonstrated by siRNA-mediated knockdown of hCUGBP1 that led to stabilization of a reporter RNA bearing the c-jun UG -rich ARE. [16]
UGU(G/A) tetranucleotides are key determinants of the binding site for xCUGBP1. A SELEX approach for the identification of artificial substrate of hCUGBP1 led to the proposition that UGU containing sequences were highly favoured for binding. [17] More recently, the reappraisal of CUGBP1 binding sites on the base of a combination of the SELEX approach and
Immunoprecipitation of the CUGBP1 containing complexes has led Graindorge et al. to propose a 15 nt motif as a key determinant of CUGBP1 binding. [18] Such a motif is found in a number of unstable mRNAs in human cells [16] suggesting that they are degraded by a CUGBP1 deadenylation dependent pathway.

In molecular biology, messenger ribonucleic acid (mRNA) is a single-stranded molecule of RNA that corresponds to the genetic sequence of a gene, and is read by a ribosome in the process of synthesizing a protein.
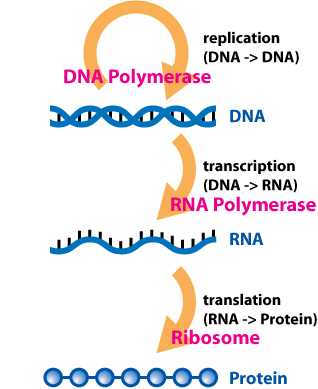
In molecular genetics, the three prime untranslated region (3′-UTR) is the section of messenger RNA (mRNA) that immediately follows the translation termination codon. The 3′-UTR often contains regulatory regions that post-transcriptionally influence gene expression.

The TATA-binding protein (TBP) is a general transcription factor that binds specifically to a DNA sequence called the TATA box. This DNA sequence is found about 30 base pairs upstream of the transcription start site in some eukaryotic gene promoters.
A trinucleotide repeat expansion, also known as a triplet repeat expansion, is the DNA mutation responsible for causing any type of disorder categorized as a trinucleotide repeat disorder. These are labelled in dynamical genetics as dynamic mutations. Triplet expansion is caused by slippage during DNA replication, also known as "copy choice" DNA replication. Due to the repetitive nature of the DNA sequence in these regions, 'loop out' structures may form during DNA replication while maintaining complementary base pairing between the parent strand and daughter strand being synthesized. If the loop out structure is formed from the sequence on the daughter strand this will result in an increase in the number of repeats. However, if the loop out structure is formed on the parent strand, a decrease in the number of repeats occurs. It appears that expansion of these repeats is more common than reduction. Generally, the larger the expansion the more likely they are to cause disease or increase the severity of disease. Other proposed mechanisms for expansion and reduction involve the interaction of RNA and DNA molecules.
An exonic splicing silencer (ESS) is a short region of an exon and is a cis-regulatory element. A set of 103 hexanucleotides known as FAS-hex3 has been shown to be abundant in ESS regions. ESSs inhibit or silence splicing of the pre-mRNA and contribute to constitutive and alternate splicing. To elicit the silencing effect, ESSs recruit proteins that will negatively affect the core splicing machinery.
Poly(A)-binding protein is an RNA-binding protein which triggers the binding of eukaryotic initiation factor 4 complex (eIF4G) directly to the poly(A) tail of mRNA which is 200-250 nucleotides long. The poly(A) tail is located on the 3' end of mRNA and was discovered by Mary Edmonds, who also characterized the poly-A polymerase enzyme that generates the poly(a) tail. The binding protein is also involved in mRNA precursors by helping polyadenylate polymerase add the poly(A) nucleotide tail to the pre-mRNA before translation. The nuclear isoform selectively binds to around 50 nucleotides and stimulates the activity of polyadenylate polymerase by increasing its affinity towards RNA. Poly(A)-binding protein is also present during stages of mRNA metabolism including nonsense-mediated decay and nucleocytoplasmic trafficking. The poly(A)-binding protein may also protect the tail from degradation and regulate mRNA production. Without these two proteins in-tandem, then the poly(A) tail would not be added and the RNA would degrade quickly.

Myotonin-protein kinase (MT-PK) also known as myotonic dystrophy protein kinase (MDPK) or dystrophia myotonica protein kinase (DMPK) is an enzyme that in humans is encoded by the DMPK gene.

Y box binding protein 1 also known as Y-box transcription factor or nuclease-sensitive element-binding protein 1 is a protein that in humans is encoded by the YBX1 gene.
Poly(A)-specific ribonuclease (PARN), also known as polyadenylate-specific ribonuclease or deadenylating nuclease (DAN), is an enzyme that in humans is encoded by the PARN gene.

CUGBP, Elav-like family member 2, also known as Etr-3 is a protein that in humans is encoded by the CELF2 gene.

Regulator of nonsense transcripts 3B is a protein that in humans is encoded by the UPF3B gene.

Serine/threonine-protein kinase MRCK alpha is an enzyme that in humans is encoded by the CDC42BPA gene.

Nesprin-2 is a protein that in humans is encoded by the SYNE2 gene. The human SYNE2 gene consists of 116 exons and encodes nesprin-2, a member of the nuclear envelope (NE) spectrin-repeat (nesprin) family. Nesprins are modular proteins with a central extended spectrin-repeat (SR) rod domain and a C-terminal Klarsicht/ANC-1/Syne homology (KASH) transmembrane domain, which acts as a NE-targeting motif. Nesprin-2 (Nesp2) binds to cytoplasmic F-actin, tethering the nucleus to the cytoskeleton and maintaining the structural integrity of the nucleus.

Muscleblind Like Splicing Regulator 1 (MBNL1) is an RNA splicing protein that in humans is encoded by the MBNL1 gene. It has a well characterized role in Myotonic dystrophy where impaired splicing disrupts muscle development and function. In addition to regulating mRNA maturation of hundreds of genes MBNL1 autoregulate alternative splicing of the MBNL1 pre-mRNA transcript. The founding member of the human MBNL family of proteins was the Drosophila Muscleblind protein.

Cellular nucleic acid-binding protein is a protein that in humans is encoded by the CNBP gene.
Dystrophia myotonica WD repeat-containing protein is a protein that in humans is encoded by the DMWD gene.

Tripartite motif-containing protein 32 is a protein that in humans is encoded by the TRIM32 gene. Since its discovery in 1995, TRIM32 has been shown to be implicated in a number of diverse biological pathways.
Maternal to zygotic transition (MZT), also known as embryonic genome activation, is the stage in embryonic development during which development comes under the exclusive control of the zygotic genome rather than the maternal (egg) genome. The egg contains stored maternal genetic material mRNA which controls embryo development until the onset of MZT. After MZT the diploid embryo takes over genetic control. This requires both zygotic genome activation (ZGA), and degradation of maternal products. This process is important because it is the first time that the new embryonic genome is utilized and the paternal and maternal genomes are used in combination. The zygotic genome now drives embryo development.
RNA-dominant diseases are characterized by deleterious mutations that typically result in degenerative disorders affecting various neurological, cardiovascular, and muscular functions. Studies have found that they arise from repetitive non-coding RNA sequences, also known as toxic RNA, which inhibit RNA-binding proteins leading to pathogenic effects. The most studied RNA-dominant diseases include, but are not limited to, myotonic dystrophy and fragile X-associated tremor/ataxia syndrome (FXTAS).
RNA-targeting small molecules represent a class of small molecules, organic compounds with traditional drug properties that can bind to RNA secondary or tertiary structures and alter translation patterns, localization, and degradation.
PDB gallery | |
|---|---|
|