
The Carboniferous is a geologic period and system of the Paleozoic that spans 60 million years from the end of the Devonian Period 358.9 Ma to the beginning of the Permian Period, 298.9 Ma. In North America, the Carboniferous is often treated as two separate geological periods, the earlier Mississippian and the later Pennsylvanian.
The Pennsylvanian is, on the ICS geologic timescale, the younger of two subperiods of the Carboniferous Period. It lasted from roughly 323.2 million years ago to 298.9 million years ago. As with most other geochronologic units, the rock beds that define the Pennsylvanian are well identified, but the exact date of the start and end are uncertain by a few hundred thousand years. The Pennsylvanian is named after the U.S. state of Pennsylvania, where the coal-producing beds of this age are widespread.
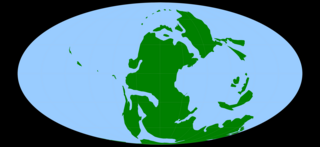
The Permian is a geologic period and stratigraphic system which spans 47 million years from the end of the Carboniferous Period 298.9 million years ago (Mya), to the beginning of the Triassic Period 251.902 Mya. It is the last period of the Paleozoic Era; the following Triassic Period belongs to the Mesozoic Era. The concept of the Permian was introduced in 1841 by geologist Sir Roderick Murchison, who named it after the region of Perm in Russia.
The PaleozoicEra is the first of three geological eras of the Phanerozoic Eon. Beginning 538.8 million years ago (Ma), it succeeds the Neoproterozoic and ends 251.9 Ma at the start of the Mesozoic Era. The Paleozoic is subdivided into six geologic periods :
The Phanerozoic is the current and the latest of the four geologic eons in the Earth's geologic time scale, covering the time period from 538.8 million years ago to the present. It is the eon during which abundant animal and plant life has proliferated, diversified and colonized various niches on the Earth's surface, beginning with the Cambrian period when animals first developed hard shells that can be clearly preserved in the fossil record. The time before the Phanerozoic, collectively called the Precambrian, is now divided into the Hadean, Archaean and Proterozoic eons.

"Labyrinthodontia" is an informal grouping of extinct predatory amphibians which were major components of ecosystems in the late Paleozoic and early Mesozoic eras. Traditionally considered a subclass of the class Amphibia, modern classification systems recognize that labyrinthodonts are not a formal natural group (clade) exclusive of other tetrapods. Instead, they consistute an evolutionary grade, ancestral to living tetrapods such as lissamphibians and amniotes. "Labyrinthodont"-grade vertebrates evolved from lobe-finned fishes in the Devonian, though a formal boundary between fish and amphibian is difficult to define at this point in time.

Diadectomorpha is a clade of large tetrapods that lived in Euramerica during the Carboniferous and Early Permian periods and in Asia during Late Permian (Wuchiapingian), They have typically been classified as advanced reptiliomorphs positioned close to, but outside of the clade Amniota, though some recent research has recovered them as the sister group to the traditional Synapsida within Amniota, based on inner ear anatomy and cladistic analyses. They include both large carnivorous and even larger herbivorous forms, some semi-aquatic and others fully terrestrial. The diadectomorphs seem to have originated during late Mississippian times, although they only became common after the Carboniferous rainforest collapse and flourished during the Late Pennsylvanian and Early Permian periods.

Joggins is a rural community located in western Cumberland County, Nova Scotia, Canada. On July 7, 2008 a 15-km length of the coast constituting the Joggins Fossil Cliffs was officially inscribed on the World Heritage List.

Arthropleura is an extinct genus of massive millipedes that lived in what is now North America and Europe around 345 to 290 million years ago, from the Viséan stage of the lower Carboniferous Period to the Sakmarian stage of the lower Permian Period. The species of the genus are the largest known land invertebrates of all time, and would have had few, if any, predators.

The late Paleozoic icehouse, also known as the Late Paleozoic Ice Age (LPIA) and formerly known as the Karoo ice age, was an ice age that began in the Late Devonian and ended in the Late Permian, occurring from 360 to 255 million years ago (Mya), and large land-based ice-sheets were then present on Earth's surface. It was the second major icehouse period of the Phanerozoic.
The Kasimovian is a geochronologic age or chronostratigraphic stage in the ICS geologic timescale. It is the third stage in the Pennsylvanian, lasting from 307 to 303.7 Ma. The Kasimovian Stage follows the Moscovian and is followed by the Gzhelian. The Kasimovian saw an extinction event which occurred around 305 mya, referred to as the Carboniferous Rainforest Collapse. It roughly corresponds to the Missourian in North American geochronology and the Stephanian in western European geochronology.

Limnoscelis was a genus of large diadectomorph tetrapods from the Late Carboniferous to early Permian of western North America. It includes two species: the type species Limnoscelis paludis from New Mexico, and Limnoscelis dynatis from Colorado, both of which are thought to have lived concurrently. No specimens of Limnoscelis are known from outside of North America. Limnoscelis was carnivorous, and likely semiaquatic, though it may have spent a significant portion of its life on land. Limnoscelis had a combination of derived amphibian and primitive reptilian features, and its placement relative to Amniota has significant implications regarding the origins of the first amniotes.

The Allegheny Group, often termed the Allegheny Formation, is a Pennsylvanian-age geological unit in the Appalachian Plateau. It is a major coal-bearing unit in the eastern United States, extending through western and central Pennsylvania, western Maryland and West Virginia, and southeastern Ohio. Fossils of fishes such as Bandringa are known from the Kittaning Formation, which is part of the Allegheny Group.
Macromerion is an extinct genus of non-mammalian synapsids, specifically Pelycosaurs, in the family Sphenacodontidae from Late Carboniferous deposits in the Czech Republic. It was named as a species of Labyrinthodon in 1875 and as its own genus in 1879.

The Medullosales is an extinct order of pteridospermous seed plants characterised by large ovules with circular cross-section and a vascularised nucellus, complex pollen-organs, stems and rachides with a dissected stele, and frond-like leaves. Their nearest still-living relatives are the cycads.
The Westphalian is a regional stage or age in the regional stratigraphy of northwest Europe, with an age between roughly 315 and 307 Ma. It is a subdivision of the Carboniferous System or Period and the regional Silesian Series. The Westphalian is named for the region of Westphalia in western Germany where strata of this age occur. The Coal Measures of England and Wales are also largely of Westphalian age, though they also extend into the succeeding Stephanian.
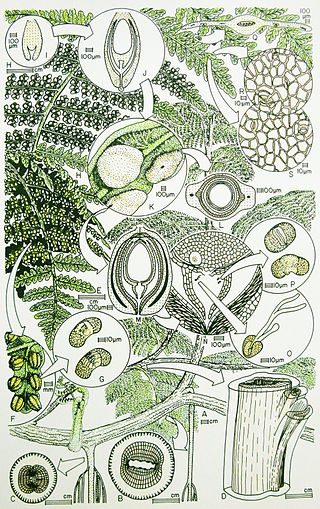
The Callistophytaceae was a family of seed ferns (pteridosperms) from the Carboniferous and Permian periods. They first appeared in late Middle Pennsylvanian (Moscovian) times, 306.5–311.7 million years ago (Ma) in the tropical coal forests of Euramerica, and became an important component of Late Pennsylvanian vegetation of clastic soils and some peat soils. The best known callistophyte was documented from Late Pennsylvanian coal ball petrifactions in North America.
Olson's Extinction was a mass extinction that occurred 273 million years ago in the late Cisuralian or early Guadalupian epoch of the Permian period, predating the much larger Permian–Triassic extinction event. The event is named after American paleontologist Everett C. Olson, who first identified the gap in fossil record indicating a sudden change between the early Permian and middle/late Permian faunas. Some authors also place a hiatus in the continental fossil record around that time, but others disagree. This event has been argued by some authors to have affected many taxa, including embryophytes, marine metazoans, and tetrapods.

Macroneuropteris is a genus of Carboniferous seed plants in the order Medullosales. The genus is best known for the species Macroneuropteris scheuchzeri, a medium-size tree that was common throughout the late Carboniferous Euramerica. Three similar species, M. macrophylla, M. britannica and M. subauriculata are also included in the genus.
This timeline of Permian research is a chronological listing of events in the history of geology and paleontology focused on the study of earth during the span of time lasting from 298.9–252.17 million years ago and the legacies of this period in the rock and fossil records.


