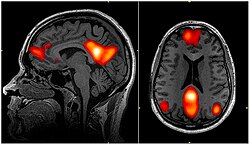
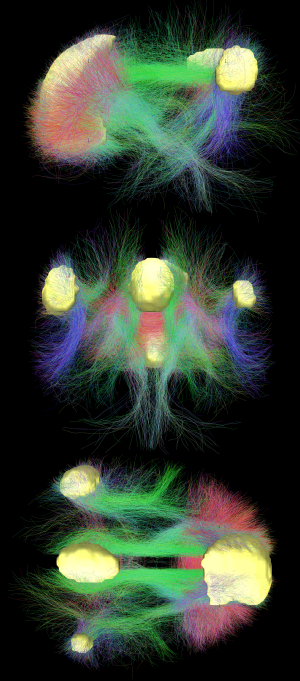
Default mode network connectivity. This image shows main regions of the default mode network (yellow) and connectivity between the regions color-coded by structural traversing direction (xyz → rgb).
In neuroscience, the default mode network (DMN), also known as the default network, default state network, or anatomically the medial frontoparietal network (M-FPN), is a large-scale brain network primarily composed of the dorsal medial prefrontal cortex, posterior cingulate cortex, precuneus and angular gyrus. It is best known for being active when a person is not focused on the outside world and the brain is at wakeful rest, such as during daydreaming and mind-wandering. It can also be active during detailed thoughts related to external task performance.[3] Other times that the DMN is active include when the individual is thinking about others, thinking about themselves, remembering the past, and planning for the future.[4][5]
The DMN was originally noticed to be deactivated in certain goal-oriented tasks and was sometimes referred to as the task-negative network,[6] in contrast with the task-positive network. This nomenclature is now widely considered misleading, because the network can be active in internal goal-oriented and conceptual cognitive tasks.[7][8][9][10] The DMN has been shown to be negatively correlated with other networks in the brain such as attention networks.[11]
Hans Berger, the inventor of the electroencephalogram, was the first to propose the idea that the brain is constantly busy. In a series of papers published in 1929, he showed that the electrical oscillations detected by his device do not cease even when the subject is at rest. However, his ideas were not taken seriously, and a general perception formed among neurologists that only when a focused activity is performed does the brain (or a part of the brain) become active.[12]
But in the 1950s, Louis Sokoloff and his colleagues noticed that metabolism in the brain stayed the same when a person went from a resting state to performing effortful math problems, suggesting active metabolism in the brain must also be happening during rest.[4] In the 1970s, David H. Ingvar and colleagues observed blood flow in the front part of the brain became the highest when a person is at rest.[4] Around the same time, intrinsic oscillatory behavior in vertebrate neurons was observed in cerebellar Purkinje cells, inferior olivary nucleus and thalamus.[13]
In the 1990s, with the advent of positron emission tomography (PET) scans, researchers began to notice that when a person is involved in perception, language, and attention tasks, the same brain areas become less active compared to passive rest, and labeled these areas as becoming "deactivated".[4]
In 1995, Bharat Biswal, a graduate student at the Medical College of Wisconsin in Milwaukee, discovered that the human sensorimotor system displayed "resting-state connectivity," exhibiting synchronicity in functional magnetic resonance imaging (fMRI) scans while not engaged in any task.[14][15]
Later, experiments by neurologistMarcus E. Raichle's lab at Washington University School of Medicine and other groups[16] showed that the brain's energy consumption is increased by less than 5% of its baseline energy consumption while performing a focused mental task. These experiments showed that the brain is constantly active with a high level of activity even when the person is not engaged in focused mental work. Research thereafter focused on finding the regions responsible for this constant background activity level.[12]
Raichle coined the term "default mode" in 2001 to describe resting state brain function;[17] the concept rapidly became a central theme in neuroscience.[18] Around this time the idea was developed that this network of brain areas is involved in internally directed thoughts and is suspended during specific goal-directed behaviors. In 2003, Greicius and colleagues examined resting state fMRI scans and looked at how correlated different sections in the brain are to each other. Their correlation maps highlighted the same areas already identified by the other researchers.[19] This was important because it demonstrated a convergence of methods all leading to the same areas being involved in the DMN. Since then other networks have been identified, such as visual, auditory, and attention networks. Some of them are often anti-correlated with the default mode network.[11]
Until the mid-2000s, researchers labeled the default mode network as the "task-negative network" because it was deactivated when participants had to perform external goal-directed tasks.[6] DMN was thought to only be active during passive rest and inactive during tasks. However, more recent studies have demonstrated the DMN to be active in certain internal goal-directed tasks such as social working memory and autobiographical tasks.[7]
Around 2007, the number of papers referencing the default mode network skyrocketed.[20] In all years prior to 2007, there were 12 papers published that referenced "default mode network" or "default network" in the title; however, between 2007 and 2014 the number increased to 1,384 papers. One reason for the increase in papers was the robust effect of finding the DMN with resting-state scans and independent component analysis (ICA).[16][21] Another reason was that the DMN could be measured with short and effortless resting-state scans, meaning they could be performed on any population including young children, clinical populations, and nonhuman primates.[4] A third reason was that the role of the DMN had been expanded to more than just a passive brain network.[4]
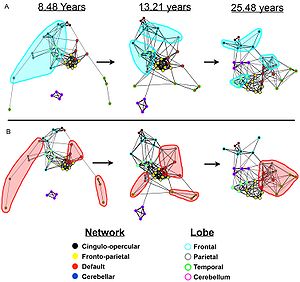
Graphs of the dynamic development of correlations between brain networks. (A) In children the regions are largely local and are organized by their physical location; the frontal regions are highlighted in light blue. (B) In adults the networks become highly correlated despite their physical distance; the default network is highlighted in light red. This result is now believed to have been confounded by artifactual processes attributable to the tendency of younger subjects to move more during image acquisition, which preferentially inflates estimates of connectivity between physically proximal regions (Power 2012, Satterthwaite 2012).
The default mode network is an interconnected and anatomically defined[4] set of brain regions. The network can be separated into hubs and subsections:
Functional hubs:[23] Information regarding the self
Posterior cingulate cortex (PCC) & precuneus: Combines bottom-up (not controlled) attention with information from memory and perception. The ventral (lower) part of PCC activates in all tasks which involve the DMN including those related to the self, related to others, remembering the past, thinking about the future, and processing concepts plus spatial navigation. The dorsal (upper) part of PCC involves involuntary awareness and arousal. The precuneus is involved in visual, sensorimotor, and attentional information.
Medial prefrontal cortex (mPFC): Decisions about self-processing such as personal information, autobiographical memories, future goals and events, and decision making regarding those personally very close such as family. The ventral (lower) part is involved in positive emotional information and internally valued reward.
Angular gyrus: Connects perception, attention, spatial cognition, and action and helps with parts of recall of episodic memories.
Dorsal medial subsystem:[23] Thinking about others
The default mode network is most commonly defined with resting state data by putting a seed in the posterior cingulate cortex and examining which other brain areas most correlate with this area.[19] The DMN can also be defined by the areas deactivated during external directed tasks compared to rest.[17]Independent component analysis (ICA) robustly finds the DMN for individuals and across groups, and has become the standard tool for mapping the default network.[16][21]
It has been shown that the default mode network exhibits the highest overlap in its structural and functional connectivity, which suggests that the structural architecture of the brain may be built in such a way that this particular network is activated by default.[1] Recent evidence from a population brain-imaging study of 10,000 UK Biobank participants further suggests that each DMN node can be decomposed into subregions with complementary structural and functional properties. It has been a widespread practice in DMN research to treat its constituent nodes to be functionally homogeneous, but the distinction between subnodes within each major DMN node has mostly been neglected. However, the close proximity of subnodes that propagate hippocampal space-time outputs and subnodes that describe the global network architecture may enable default functions, such as autobiographical recall or internally-orientated thinking.[25]
In the infant's brain, there is limited evidence of the default network, but default network connectivity is more consistent in children aged 9–12 years, suggesting that the default network undergoes developmental change.[11]
Functional connectivity analysis in monkeys shows a similar network of regions to the default mode network seen in humans.[4] The PCC is also a key hub in monkeys; however, the mPFC is smaller and less well connected to other brain regions, largely because human's mPFC is much larger and well developed.[4]
Diffusion MRI imaging shows white matter tracts connecting different areas of the DMN together.[20] The structural connections found from diffusion MRI imaging and the functional correlations from resting state fMRI show the highest level of overlap and agreement within the DMN areas.[1] This provides evidence that neurons in the DMN regions are linked to each other through large tracts of axons and this causes activity in these areas to be correlated with one another. From the point of view of effective connectivity, many studies have attempted to shed some light using dynamic causal modeling, with inconsistent results. However, directionality from the medial prefrontal cortex towards the posterior cingulate gyrus seems confirmed in multiple studies, and the inconsistent results appear to be related to small sample size analysis.[26]
Function
The default mode network is thought to be involved in several different functions:
It is potentially the neurological basis for the self:[20]
Autobiographical information: Memories of collection of events and facts about one's self
Self-reference: Referring to traits and descriptions of one's self
Emotion of one's self: Reflecting about one's own emotional state
The default mode network is active during passive rest and mind-wandering[4] which usually involves thinking about others, thinking about one's self, remembering the past, and envisioning the future rather than the task being performed.[20] Recent work, however, has challenged a specific mapping between the default mode network and mind-wandering, given that the system is important in maintaining detailed representations of task information during working memory encoding.[29]Electrocorticography studies (which involve placing electrodes on the surface of a subject's cerebral cortex) have shown the default mode network becomes activated within a fraction of a second after participants finish a task.[30] Additionally, during attention demanding tasks, sufficient deactivation of the default mode network at the time of memory encoding has been shown to result in more successful long-term memory consolidation.[31]
Studies have shown that when people watch a movie,[32] listen to a story,[33][34] or read a story,[35] their DMNs are highly correlated with each other. DMNs are not correlated if the stories are scrambled or are in a language the person does not understand, suggesting that the network is highly involved in the comprehension and the subsequent memory formation of that story.[34] The DMN is shown to even be correlated if the same story is presented to different people in different languages,[36] further suggesting the DMN is truly involved in the comprehension aspect of the story and not the auditory or language aspect.
The default mode network is deactivated during some external goal-oriented tasks such as visual attention or cognitive working memory tasks.[6] However, with internal goal-oriented tasks, such as social working memory or autobiographical tasks, the DMN is positively activated with the task and correlates with other networks such as the network involved in executive function.[7] Regions of the DMN are also activated during cognitively demanding tasks that require higher-order conceptual representations.[9] The DMN shows higher activation when behavioral responses are stable, and this activation is independent of self-reported mind wandering.[37]
Tsoukalas (2017) links theory of mind to immobilization, and suggests that the default network is activated by the immobilization inherent in the testing procedure (the patient is strapped supine on a stretcher and inserted by a narrow tunnel into a massive metallic structure). This procedure creates a sense of entrapment and, not surprisingly, the most commonly reported side-effect is claustrophobia.[38]
Gabrielle et al. (2019) suggests that the DMN is related to the perception of beauty, in which the network becomes activated in a generalized way to aesthetically moving domains such as artworks, landscapes, and architecture. This would explain a deep inner feeling of pleasure related to aesthetics, interconnected with the sense of personal identity, due to the network functions related to the self.[39]
People with Alzheimer's disease show a reduction in glucose (energy use) within the areas of the default mode network.[4] These reductions start off as slight decreases in patients with mild symptoms and continue to large reductions in those with severe symptoms. Surprisingly, disruptions in the DMN begin even before individuals show signs of Alzheimer's disease.[4] Plots of the peptide amyloid-beta, which is thought to cause Alzheimer's disease, show the buildup of the peptide is within the DMN.[4] This prompted Randy Buckner and colleagues to propose the high metabolic rate from continuous activation of DMN causes more amyloid-beta peptide to accumulate in these DMN areas.[4] These amyloid-beta peptides disrupt the DMN and because the DMN is heavily involved in memory formation and retrieval, this disruption leads to the symptoms of Alzheimer's disease.
DMN is thought to be disrupted in individuals with autism spectrum disorder.[4][42] These individuals are impaired in social interaction and communication which are tasks central to this network. Studies have shown worse connections between areas of the DMN in individuals with autism, especially between the mPFC (involved in thinking about the self and others) and the PCC (the central core of the DMN).[43][44] The more severe the autism, the less connected these areas are to each other.[43][44] It is not clear if this is a cause or a result of autism, or if a third factor is causing both (confounding).
Although it is not clear whether the DMN connectivity is increased or decreased in psychotic bipolar disorder and schizophrenia, several genes correlated with altered DMN connectivity are also risk genes for mood and psychosis disorders.[45]
Rumination, one of the main symptoms of major depressive disorder, is associated with increased DMN connectivity and dominance over other networks during rest.[46][47] Such DMN hyperconnectivity has been observed in first-episode depression[48] and chronic pain.[49] Altered DMN connectivity may change the way a person perceives events and their social and moral reasoning, thus increasing their susceptibility to depressive symptoms.[50]
Lower connectivity between brain regions was found across the default network in people who have experienced long-term trauma, such as childhood abuse or neglect, and is associated with dysfunctional attachment patterns. Among people experiencing PTSD, lower activation was found in the posterior cingulate gyrus compared to controls, and severe PTSD was characterized by lower connectivity within the DMN.[40][51]
Adults and children with ADHD show reduced anticorrelation between the DMN and other brain networks.[52][53] The cause may be a lag in brain maturation.[54] More generally, competing activation between the DMN and other networks during memory encoding may result in poor long-term memory consolidation, which is a symptom of not only ADHD but also depression, anxiety, autism, and schizophrenia.[31]
Modulation
The default mode network (DMN) may be modulated by the following interventions and processes:
Acupuncture – Deactivation of the limbic brain areas and the DMN.[55] It has been suggested that this is due to the pain response.[56]
Antidepressants – Abnormalities in DMN connectivity are reduced following treatment with antidepressant medications in PTSD.[57]
Attention Training Technique - Research shows that even a single session of Attention Training Technique changes functional connectivity of the DMN.[58]
Deep brain stimulation – Alterations in brain activity with deep brain stimulation may be used to balance resting state networks.[59]
Physical Activityand Exercise – Physical Activity, and more likely Aerobic Training, may alter the DMN. In addition, sports experts are showing networks differences, notably of the DMN.[63][64][65]
Psychedelic drugs – Reduced blood flow to the PCC and mPFC was observed under the administration of psilocybin. These two areas are considered to be the main nodes of the DMN.[66] One study on the effects of LSD demonstrated that the drug desynchronizes brain activity within the DMN; the activity of the brain regions that constitute the DMN becomes less correlated.[67]
Psychotherapy – In PTSD, the abnormalities in the default mode network normalize in individuals who respond to psychotherapy interventions.[68][57]
Sleep deprivation – Functional connectivity between nodes of the DMN in their resting-state is usually strong, but sleep deprivation results in a decrease in connectivity within the DMN.[69] Recent studies suggest a decrease in connectivity between the DMN and the task-positive network as a result of sleep loss.[70]
Stage N3 of NREM sleep – Further decrease in connectivity between the PCC and MPFC.[71]
Criticism
Some have argued the brain areas in the default mode network only show up together because of the vascular coupling of large arteries and veins in the brain near these areas, not because these areas are actually functionally connected to each other. Support for this argument comes from studies that show changing in breathing alters oxygen levels in the blood which in turn affects DMN the most.[4] These studies however do not explain why the DMN can also be identified using PET scans by measuring glucose metabolism which is independent of vascular coupling[4] and in electrocorticography studies[72] measuring electrical activity on the surface of the brain, and in MEG by measuring magnetic fields associated with electrophysiological brain activity that bypasses the hemodynamic response.[73]
The idea of a "default network" is not universally accepted.[74] In 2007 the concept of the default mode was criticized as not being useful for understanding brain function, on the grounds that a simpler hypothesis is that a resting brain actually does more processing than a brain doing certain "demanding" tasks, and that there is no special significance to the intrinsic activity of the resting brain.[75]
Nomenclature
The default mode network has also been called the language network, semantic system, or limbic network.[10] Even though the dichotomy is misleading,[7] the term task-negative network is still sometimes used to contrast it against other more externally-oriented brain networks.[53]
In 2019, Uddin et al. proposed that medial frontoparietal network (M-FPN) be used as a standard anatomical name for this network.[10]
In neuroanatomy, the precuneus is the portion of the superior parietal lobule on the medial surface of each brain hemisphere. It is located in front of the cuneus. The precuneus is bounded in front by the marginal branch of the cingulate sulcus, at the rear by the parieto-occipital sulcus, and underneath by the subparietal sulcus. It is involved with episodic memory, visuospatial processing, reflections upon self, and aspects of consciousness.
Brodmann area 9, or BA9, refers to a cytoarchitecturally defined portion of the frontal cortex in the brain of humans and other primates. Its cytoarchitecture is referred to as granular due to the concentration of granule cells in layer IV. It contributes to the dorsolateral and medial prefrontal cortex.
The angular gyrus is a region of the brain lying mainly in the posteroinferior region of the parietal lobe, occupying the posterior part of the inferior parietal lobule. It represents the Brodmann area 39.
The posterior cingulate cortex (PCC) is the caudal part of the cingulate cortex, located posterior to the anterior cingulate cortex. This is the upper part of the "limbic lobe". The cingulate cortex is made up of an area around the midline of the brain. Surrounding areas include the retrosplenial cortex and the precuneus.
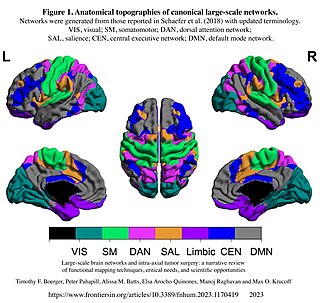
The dorsal attention network (DAN), also known anatomically as the dorsal frontoparietal network (D-FPN), is a large-scale brain network of the human brain that is primarily composed of the intraparietal sulcus (IPS) and frontal eye fields (FEF). It is named and most known for its role in voluntary orienting of visuospatial attention.
Connectomics is the production and study of connectomes: comprehensive maps of connections within an organism's nervous system. More generally, it can be thought of as the study of neuronal wiring diagrams with a focus on how structural connectivity, individual synapses, cellular morphology, and cellular ultrastructure contribute to the make up of a network. The nervous system is a network made of billions of connections and these connections are responsible for our thoughts, emotions, actions, memories, function and dysfunction. Therefore, the study of connectomics aims to advance our understanding of mental health and cognition by understanding how cells in the nervous system are connected and communicate. Because these structures are extremely complex, methods within this field use a high-throughput application of functional and structural neural imaging, most commonly magnetic resonance imaging (MRI), electron microscopy, and histological techniques in order to increase the speed, efficiency, and resolution of these nervous system maps. To date, tens of large scale datasets have been collected spanning the nervous system including the various areas of cortex, cerebellum, the retina, the peripheral nervous system and neuromuscular junctions.
Marcus E. Raichle is an American neurologist at the Washington University School of Medicine in Saint Louis, Missouri. He is a professor in the Department of Radiology with joint appointments in Neurology, Neurobiology and Biomedical Engineering. His research over the past 40 years has focused on the nature of functional brain imaging signals arising from PET and fMRI and the application of these techniques to the study of the human brain in health and disease. He received the Kavli Prize in Neuroscience “for the discovery of specialized brain networks for memory and cognition", together with Brenda Milner and John O’Keefe in 2014.
Resting state fMRI is a method of functional magnetic resonance imaging (fMRI) that is used in brain mapping to evaluate regional interactions that occur in a resting or task-negative state, when an explicit task is not being performed. A number of resting-state brain networks have been identified, one of which is the default mode network. These brain networks are observed through changes in blood flow in the brain which creates what is referred to as a blood-oxygen-level dependent (BOLD) signal that can be measured using fMRI.
The biological basis of personality is a collection of brain systems and mechanisms that underlie human personality. Human neurobiology, especially as it relates to complex traits and behaviors, is not well understood, but research into the neuroanatomical and functional underpinnings of personality are an active field of research. Animal models of behavior, molecular biology, and brain imaging techniques have provided some insight into human personality, especially trait theories.
The dorsal nexus is an area within the dorsal medial prefrontal cortex that serves as an intersection point for multiple brain networks. Research suggests it plays a role in the maintenance and manipulation of information, as well as supporting the control of cognitive functions such as behavior, memory, and conflict resolution. Abnormally increased connectivity between these networks through the dorsal nexus has been associated with certain types of depression. The activity generated by this abnormally high level of connectivity during a depressive state can be identified through magnetic resonance imaging (MRI) and positron emission tomography (PET).
Dynamic functional connectivity (DFC) refers to the observed phenomenon that functional connectivity changes over a short time. Dynamic functional connectivity is a recent expansion on traditional functional connectivity analysis which typically assumes that functional networks are static in time. DFC is related to a variety of different neurological disorders, and has been suggested to be a more accurate representation of functional brain networks. The primary tool for analyzing DFC is fMRI, but DFC has also been observed with several other mediums. DFC is a recent development within the field of functional neuroimaging whose discovery was motivated by the observation of temporal variability in the rising field of steady state connectivity research.
Large-scale brain networks are collections of widespread brain regions showing functional connectivity by statistical analysis of the fMRI BOLD signal or other recording methods such as EEG, PET and MEG. An emerging paradigm in neuroscience is that cognitive tasks are performed not by individual brain regions working in isolation but by networks consisting of several discrete brain regions that are said to be "functionally connected". Functional connectivity networks may be found using algorithms such as cluster analysis, spatial independent component analysis (ICA), seed based, and others. Synchronized brain regions may also be identified using long-range synchronization of the EEG, MEG, or other dynamic brain signals.
An identity disturbance is a deficiency or inability to maintain one or more major components of identity. These components include a sense of continuity over time; emotional commitment to representations of self, role relationships, core values and self-standards; development of a meaningful world view; and recognition of one's place in the world.
The salience network (SN), also known anatomically as the midcingulo-insular network (M-CIN) or ventral attention network, is a large scale network of the human brain that is primarily composed of the anterior insula (AI) and dorsal anterior cingulate cortex (dACC). It is involved in detecting and filtering salient stimuli, as well as in recruiting relevant functional networks. Together with its interconnected brain networks, the SN contributes to a variety of complex functions, including communication, social behavior, and self-awareness through the integration of sensory, emotional, and cognitive information.
The sensorimotor network (SMN), also known as somatomotor network, is a large-scale brain network that primarily includes somatosensory and motor regions and extends to the supplementary motor areas (SMA). The auditory cortex may also be included. The SMN is activated during motor tasks, such as finger tapping, indicating that the network readies the brain when performing and coordinating motor tasks.
Social cognitive neuroscience is the scientific study of the biological processes underpinning social cognition. Specifically, it uses the tools of neuroscience to study "the mental mechanisms that create, frame, regulate, and respond to our experience of the social world". Social cognitive neuroscience uses the epistemological foundations of cognitive neuroscience, and is closely related to social neuroscience. Social cognitive neuroscience employs human neuroimaging, typically using functional magnetic resonance imaging (fMRI). Human brain stimulation techniques such as transcranial magnetic stimulation and transcranial direct-current stimulation are also used. In nonhuman animals, direct electrophysiological recordings and electrical stimulation of single cells and neuronal populations are utilized for investigating lower-level social cognitive processes.
Network neuroscience is an approach to understanding the structure and function of the human brain through an approach of network science, through the paradigm of graph theory. A network is a connection of many brain regions that interact with each other to give rise to a particular function. Network Neuroscience is a broad field that studies the brain in an integrative way by recording, analyzing, and mapping the brain in various ways. The field studies the brain at multiple scales of analysis to ultimately explain brain systems, behavior, and dysfunction of behavior in psychiatric and neurological diseases. Network neuroscience provides an important theoretical base for understanding neurobiological systems at multiple scales of analysis.
The frontoparietal network (FPN), generally also known as the central executive network (CEN) or, more specifically, the lateral frontoparietal network (L-FPN), is a large-scale brain network primarily composed of the dorsolateral prefrontal cortex and posterior parietal cortex, around the intraparietal sulcus. It is involved in sustained attention, complex problem-solving and working memory.
Functional MRI imaging methods have allowed researchers to combine neurocognitive testing with structural neuroanatomical measures, take into consideration both cognitive and affective paradigms, and subsequently create computer-aided diagnosis techniques and algorithms. Functional MRI has several benefits, such as its non-invasive quality, relatively high spatial resolution, and decent temporal resolution. One particular method used in recent research is resting-state functional magnetic resonance imaging, rs-fMRI. fMRI imaging has been applied to numerous behavioral studies for schizophrenia, the findings of which have hinted toward potential brain regions that govern key characteristics in cognition and affect.
Susan Whitfield-Gabrieli is an American scientist, psychologist/neuroscientist, academic and researcher. She is a professor of psychology, the Founding Director of the Biomedical Imaging Center at Northeastern University, Researcher in the Department of Psychiatry at Massachusetts General Hospital, Harvard Medical School and a Research Affiliate of McGovern Institute for Brain Research at Massachusetts Institute of Technology.
↑ Biswal, B; Yetkin, F. Z.; Haughton, V. M.; Hyde, J. S. (1995). "Functional connectivity in the motor cortex of resting human brain using echoplanar MRI". Magn Reson Med. 34 (4): 537–541. doi:10.1002/mrm.1910340409. PMID8524021. S2CID775793.
1 2 De Luca, M; Beckmann, CF; De Stefano, N; Matthews, PM; Smith, SM (15 February 2006). "fMRI resting state networks define distinct modes of long-distance interactions in the human brain". NeuroImage. 29 (4): 1359–1367. doi:10.1016/j.neuroimage.2005.08.035. PMID16260155. S2CID16193549.
↑ Silchenko, Alexander N.; Hoffstaedter, Felix; Eickhoff, Simon B. (2023). "Impact of sample size and regression of tissue-specific signals on effective connectivity within the core default mode network". Human Brain Mapping.
↑ Vigneshwaran, S; Mahanand, B. S.; Suresh, S; Sundararajan, N (2017). "Identifying differences in brain activities and an accurate detection of autism spectrum disorder using resting state functional-magnetic resonance imaging: A spatial filtering approach". Medical Image Analysis. 35: 375–389. doi:10.1016/j.media.2016.08.003. PMID27585835. S2CID4922560.
↑ Carhart-Harris, Robin L.; Muthukumaraswamy, Suresh; Roseman, Leor; Kaelen, Mendel; Droog, Wouter; Murphy, Kevin; Tagliazucchi, Enzo; Schenberg, Eduardo E.; Nest, Timothy; Orban, Csaba; Leech, Robert; Williams, Luke T.; Williams, Tim M.; Bolstridge, Mark; Sessa, Ben; McGonigle, John; Sereno, Martin I.; Nichols, David; Hellyer, Peter J.; Hobden, Peter; Evans, John; Singh, Krish D.; Wise, Richard G.; Curran, H. Valerie; Feilding, Amanda; Nutt, David J. (26 April 2016). "Neural correlates of the LSD experience revealed by multimodal neuroimaging". Proceedings of the National Academy of Sciences. 113 (17): 4853–4858. Bibcode:2016PNAS..113.4853C. doi:10.1073/pnas.1518377113. PMC4855588. PMID27071089.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.