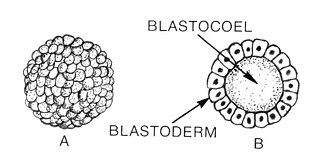
Blastulation is the stage in early animal embryonic development that produces the blastula. In mammalian development the blastula develops into the blastocyst with a differentiated inner cell mass and an outer trophectoderm. The blastula is a hollow sphere of cells known as blastomeres surrounding an inner fluid-filled cavity called the blastocoel. Embryonic development begins with a sperm fertilizing an egg cell to become a zygote, which undergoes many cleavages to develop into a ball of cells called a morula. Only when the blastocoel is formed does the early embryo become a blastula. The blastula precedes the formation of the gastrula in which the germ layers of the embryo form.
A maternal effect is a situation where the phenotype of an organism is determined not only by the environment it experiences and its genotype, but also by the environment and genotype of its mother. In genetics, maternal effects occur when an organism shows the phenotype expected from the genotype of the mother, irrespective of its own genotype, often due to the mother supplying messenger RNA or proteins to the egg. Maternal effects can also be caused by the maternal environment independent of genotype, sometimes controlling the size, sex, or behaviour of the offspring. These adaptive maternal effects lead to phenotypes of offspring that increase their fitness. Further, it introduces the concept of phenotypic plasticity, an important evolutionary concept. It has been proposed that maternal effects are important for the evolution of adaptive responses to environmental heterogeneity.

A coenocyte is a multinucleate cell which can result from multiple nuclear divisions without their accompanying cytokinesis, in contrast to a syncytium, which results from cellular aggregation followed by dissolution of the cell membranes inside the mass. The word syncytium in animal embryology is used to refer to the coenocytic blastoderm of invertebrates. A coenocytic colony is referred to as a coenobium, and most coenobia are composed of a distinct number of cells, often as a multiple of two.
Segmentation in biology is the division of some animal and plant body plans into a linear series of repetitive segments that may or may not be interconnected to each other. This article focuses on the segmentation of animal body plans, specifically using the examples of the taxa Arthropoda, Chordata, and Annelida. These three groups form segments by using a "growth zone" to direct and define the segments. While all three have a generally segmented body plan and use a growth zone, they use different mechanisms for generating this patterning. Even within these groups, different organisms have different mechanisms for segmenting the body. Segmentation of the body plan is important for allowing free movement and development of certain body parts. It also allows for regeneration in specific individuals.

A morphogen is a substance whose non-uniform distribution governs the pattern of tissue development in the process of morphogenesis or pattern formation, one of the core processes of developmental biology, establishing positions of the various specialized cell types within a tissue. More specifically, a morphogen is a signaling molecule that acts directly on cells to produce specific cellular responses depending on its local concentration.
Hox genes, a subset of homeobox genes, are a group of related genes that specify regions of the body plan of an embryo along the head-tail axis of animals. Hox proteins encode and specify the characteristics of 'position', ensuring that the correct structures form in the correct places of the body. For example, Hox genes in insects specify which appendages form on a segment, and Hox genes in vertebrates specify the types and shape of vertebrae that will form. In segmented animals, Hox proteins thus confer segmental or positional identity, but do not form the actual segments themselves.
Compartments can be simply defined as separate, different, adjacent cell populations, which upon juxtaposition, create a lineage boundary. This boundary prevents cell movement from cells from different lineages across this barrier, restricting them to their compartment. Subdivisions are established by morphogen gradients and maintained by local cell-cell interactions, providing functional units with domains of different regulatory genes, which give rise to distinct fates. Compartment boundaries are found across species. In the hindbrain of vertebrate embryos, rhombomeres are compartments of common lineage outlined by expression of Hox genes. In invertebrates, the wing imaginal disc of Drosophila provides an excellent model for the study of compartments. Although other tissues, such as the abdomen, and even other imaginal discs are compartmentalized, much of our understanding of key concepts and molecular mechanisms involved in compartment boundaries has been derived from experimentation in the wing disc of the fruit fly.

Ultrabithorax (Ubx) is a homeobox gene found in insects, and is used in the regulation of patterning in morphogenesis. There are many possible products of this gene, which function as transcription factors. Ubx is used in the specification of serially homologous structures, and is used at many levels of developmental hierarchies. In Drosophila melanogaster it is expressed in the third thoracic (T3) and first abdominal (A1) segments and represses wing formation. The Ubx gene regulates the decisions regarding the number of wings and legs the adult flies will have. The developmental role of the Ubx gene is determined by the splicing of its product, which takes place after translation of the gene. The specific splice factors of a particular cell allow the specific regulation of the developmental fate of that cell, by making different splice variants of transcription factors. In D. melanogaster, at least six different isoforms of Ubx exist.

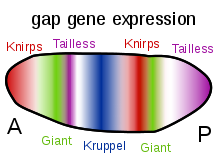
Krüppel is a gap gene in Drosophila melanogaster, located on the 2R chromosome, which encodes a zinc finger C2H2 transcription factor. Gap genes work together to establish the anterior-posterior segment patterning of the insect through regulation of the transcription factor encoding pair rule genes. These genes in turn regulate segment polarity genes. Krüppel means "cripple" in German, named for the crippled appearance of mutant larvae, who have failed to develop proper thoracic and anterior segments in the abdominal region. Mutants can also have abdominal mirror duplications.

A gap gene is a type of gene involved in the development of the segmented embryos of some arthropods. Gap genes are defined by the effect of a mutation in that gene, which causes the loss of contiguous body segments, resembling a gap in the normal body plan. Each gap gene, therefore, is necessary for the development of a section of the organism.
Decapentaplegic (Dpp) is a key morphogen involved in the development of the fruit fly Drosophila melanogaster and is the first validated secreted morphogen. It is known to be necessary for the correct patterning and development of the early Drosophila embryo and the fifteen imaginal discs, which are tissues that will become limbs and other organs and structures in the adult fly. It has also been suggested that Dpp plays a role in regulating the growth and size of tissues. Flies with mutations in decapentaplegic fail to form these structures correctly, hence the name. Dpp is the Drosophila homolog of the vertebrate bone morphogenetic proteins (BMPs), which are members of the TGF-β superfamily, a class of proteins that are often associated with their own specific signaling pathway. Studies of Dpp in Drosophila have led to greater understanding of the function and importance of their homologs in vertebrates like humans.
In the field of developmental biology, regional differentiation is the process by which different areas are identified in the development of the early embryo. The process by which the cells become specified differs between organisms.

A pair-rule gene is a type of gene involved in the development of the segmented embryos of insects. Pair-rule genes are expressed as a result of differing concentrations of gap gene proteins, which encode transcription factors controlling pair-rule gene expression. Pair-rule genes are defined by the effect of a mutation in that gene, which causes the loss of the normal developmental pattern in alternating segments.
Orthodenticle (otd) is a homeobox gene found in Drosophila that regulates the development of anterior patterning, with particular involvement in the central nervous system function and eye development. It is located on the X chromosome. The gene is an ortholog of the human OTX1/OTX2 gene.
A segmentation gene is a generic term for a gene whose function is to specify tissue pattern in each repeated unit of a segmented organism. Animals are constructed of segments; however, Drosophila segments also contain subdivided compartments. There are five gene classes which each contribute to the segmentation and development of the embryonic drosophila. These five gene classes include the coordinate gene, gap gene, pair-rule gene, segment polarity gene, and homeotic gene. In embryonic drosophila, the pair-rule gene defines odd-skipped and even-skipped genes as parasegments, showing 7 stripes in the embryo. In the next gene class, segment polarity gene, individual segments each have their own anterior and posterior pole, resulting in 14 segments. In the fruit fly Drosophila melanogaster, segment polarity genes help to define the anterior and posterior polarities within each embryonic parasegment by regulating the transmission of signals via the Wnt signaling pathway and Hedgehog signaling pathway. Segment polarity genes are expressed in the embryo following expression of the gap genes and pair-rule genes. The most commonly cited examples of these genes are engrailed and gooseberry in Drosophila melanogaster. The segment polarity is the last step in embryonic development and a repeated pattern where each half of each segment is deleted and a mirror-image is duplicated and reversed to replace that half segment; thus, forming a pattern element.

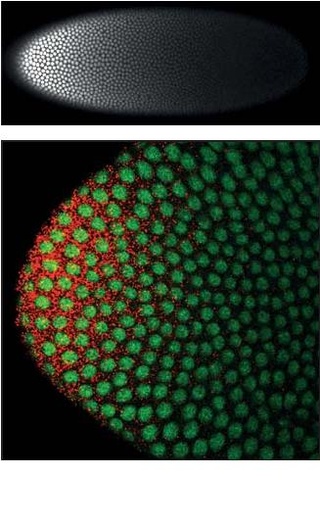
Germ-band extension is a morphological process widely studied in Drosophila melanogaster in which the germ-band, which develops into the segmented trunk of the embryo, approximately doubles in length along the anterior-posterior axis while subsequently narrowing along the dorsal-ventral axis.
Staufen is a protein product of a maternally expressed gene first identified in Drosophila melanogaster. The protein has been implicated in helping regulate genes important in determination of gradients that set up the anterior posterior axis such as bicoid and oskar. Staufen proteins, abbreviated Stau, are necessary for cell localization during the oogenesis and zygotic development. It is involved in targeting of the messenger RNA encoding these genes to the correct pole of the egg cell.
The Cdx gene family, also called caudal genes, are a group of genes found in many animal genomes. Cdx genes contain a homeobox DNA sequence and code for proteins that act as transcription factors. The gene after which the gene family is named is the caudal or cad gene of the fruitfly Drosophila melanogaster. The human genome has three Cdx genes, called CDX1, CDX2 and CDX4. The zebrafish has no cdx2 gene, but two copies of cdx1 and one copy of cdx4. The Cdx gene in the nematode Caenorhabditis elegans is called pal-1.
Homeotic protein bicoid is encoded by the bcd maternal effect gene in Drosophilia. Homeotic protein bicoid concentration gradient patterns the anterior-posterior (A-P) axis during Drosophila embryogenesis. Bicoid was the first protein demonstrated to act as a morphogen. Although bicoid is important for the development of Drosophila and other higher dipterans, it is absent from most other insects, where its role is accomplished by other genes.
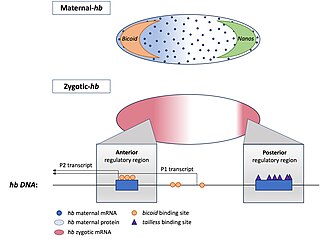
Hunchback is a maternal effect and zygotic gene expressed in the embryos of the fruit fly Drosophila melanogaster. In maternal effect genes, the RNA or protein from the mother’s gene is deposited into the oocyte or embryo before the embryo can express its own zygotic genes.