
Bacterial conjugation is the transfer of genetic material between bacterial cells by direct cell-to-cell contact or by a bridge-like connection between two cells. This takes place through a pilus. It is a parasexual mode of reproduction in bacteria.

A pilus is a hair-like appendage found on the surface of many bacteria and archaea. The terms pilus and fimbria can be used interchangeably, although some researchers reserve the term pilus for the appendage required for bacterial conjugation. All conjugative pili are primarily composed of pilin – fibrous proteins, which are oligomeric.
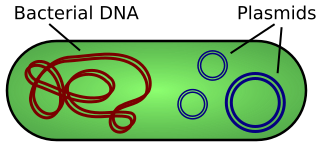
A plasmid is a small, extrachromosomal DNA molecule within a cell that is physically separated from chromosomal DNA and can replicate independently. They are most commonly found as small circular, double-stranded DNA molecules in bacteria; however, plasmids are sometimes present in archaea and eukaryotic organisms. In nature, plasmids often carry genes that benefit the survival of the organism and confer selective advantage such as antibiotic resistance. While chromosomes are large and contain all the essential genetic information for living under normal conditions, plasmids are usually very small and contain only additional genes that may be useful in certain situations or conditions. Artificial plasmids are widely used as vectors in molecular cloning, serving to drive the replication of recombinant DNA sequences within host organisms. In the laboratory, plasmids may be introduced into a cell via transformation. Synthetic plasmids are available for procurement over the internet.

A high-frequency recombination cell is a bacterium with a conjugative plasmid integrated into its chromosomal DNA. The integration of the plasmid into the cell's chromosome is through homologous recombination. A conjugative plasmid capable of chromosome integration is also called an episome. When conjugation occurs, Hfr cells are very efficient in delivering chromosomal genes of the cell into recipient F− cells, which lack the episome.

Agrobacterium radiobacter is the causal agent of crown gall disease in over 140 species of eudicots. It is a rod-shaped, Gram-negative soil bacterium. Symptoms are caused by the insertion of a small segment of DNA, from a plasmid into the plant cell, which is incorporated at a semi-random location into the plant genome. Plant genomes can be engineered by use of Agrobacterium for the delivery of sequences hosted in T-DNA binary vectors.

Transduction is the process by which foreign DNA is introduced into a cell by a virus or viral vector. An example is the viral transfer of DNA from one bacterium to another and hence an example of horizontal gene transfer. Transduction does not require physical contact between the cell donating the DNA and the cell receiving the DNA, and it is DNase resistant. Transduction is a common tool used by molecular biologists to stably introduce a foreign gene into a host cell's genome.
Genetics, a discipline of biology, is the science of heredity and variation in living organisms.

A tumour inducing (Ti) plasmid is a plasmid found in pathogenic species of Agrobacterium, including A. tumefaciens, A. rhizogenes, A. rubi and A. vitis.
Fosmids are similar to cosmids but are based on the bacterial F-plasmid. The cloning vector is limited, as a host can only contain one fosmid molecule. Fosmids can hold DNA inserts of up to 40 kb in size; often the source of the insert is random genomic DNA. A fosmid library is prepared by extracting the genomic DNA from the target organism and cloning it into the fosmid vector. The ligation mix is then packaged into phage particles and the DNA is transfected into the bacterial host. Bacterial clones propagate the fosmid library. The low copy number offers higher stability than vectors with relatively higher copy numbers, including cosmids. Fosmids may be useful for constructing stable libraries from complex genomes. Fosmids have high structural stability and have been found to maintain human DNA effectively even after 100 generations of bacterial growth. Fosmid clones were used to help assess the accuracy of the Public Human Genome Sequence.
Bacteriophage MS2, commonly called MS2, is an icosahedral, positive-sense single-stranded RNA virus that infects the bacterium Escherichia coli and other members of the Enterobacteriaceae. MS2 is a member of a family of closely related bacterial viruses that includes bacteriophage f2, bacteriophage Qβ, R17, and GA.

Prokaryotic DNA Replication is the process by which a prokaryote duplicates its DNA into another copy that is passed on to daughter cells. Although it is often studied in the model organism E. coli, other bacteria show many similarities. Replication is bi-directional and originates at a single origin of replication (OriC). It consists of three steps: Initiation, elongation, and termination.
An origin of transfer (oriT) is a short sequence ranging from 40-500 base pairs in length that is necessary for the transfer of DNA from a gram-negative bacterial donor to recipient during bacterial conjugation. The transfer of DNA is a critical component for antimicrobial resistance within bacterial cells and the oriT structure and mechanism within plasmid DNA is complementary to its function in bacterial conjugation. The first oriT to be identified and cloned was on the RK2 (IncP) conjugative plasmid, which was done by Guiney and Helinski in 1979.
The relaxosome is the complex of proteins that facilitates plasmid transfer during bacterial conjugation. The proteins are encoded by the tra operon on a fertility plasmid in the region near the origin of transfer, oriT. The most important of these proteins is relaxase, which is responsible for beginning the conjugation process by cutting at the nic site via transesterification. This nicking results in a DNA-Protein complex with the relaxosome bound to a single strand of the plasmid DNA and an exposed 3' hydroxyl group. Relaxase also unwinds the plasmid being conjugated with its helicase properties. The relaxosome interacts with integration host factors within the oriT.
Transfer genes or tra genes, are some genes necessary for non-sexual transfer of genetic material in both gram-positive and gram-negative bacteria. The tra locus includes the pilin gene and regulatory genes, which together form pili on the cell surface, polymeric proteins that can attach themselves to the surface of F-bacteria and initiate the conjugation. The existence of the tra region of a plasmid genome was first discovered in 1979 by David H. Figurski and Donald R. Helinski In the course of their work, Figurski and Helinski also discovered a second key fact about the tra region – that it can act in trans to the mobilization marker which it affects.
In molecular cloning, a vector is any particle used as a vehicle to artificially carry a foreign nucleic sequence – usually DNA – into another cell, where it can be replicated and/or expressed. A vector containing foreign DNA is termed recombinant DNA. The four major types of vectors are plasmids, viral vectors, cosmids, and artificial chromosomes. Of these, the most commonly used vectors are plasmids. Common to all engineered vectors are an origin of replication, a multicloning site, and a selectable marker.

A toxin-antitoxin system consists of a "toxin" and a corresponding "antitoxin", usually encoded by closely linked genes. The toxin is usually a protein while the antitoxin can be a protein or an RNA. Toxin-antitoxin systems are widely distributed in prokaryotes, and organisms often have them in multiple copies. When these systems are contained on plasmids – transferable genetic elements – they ensure that only the daughter cells that inherit the plasmid survive after cell division. If the plasmid is absent in a daughter cell, the unstable antitoxin is degraded and the stable toxic protein kills the new cell; this is known as 'post-segregational killing' (PSK).
Plasmid-mediated resistance is the transfer of antibiotic resistance genes which are carried on plasmids. Plasmids possess mechanisms that ensure their independent replication as well as those that regulate their replication number and guarantee stable inheritance during cell division. By the conjugation process, they can stimulate lateral transfer between bacteria from various genera and kingdoms. Numerous plasmids contain addiction-inducing systems that are typically based on toxin-antitoxin factors and capable of killing daughter cells that don't inherit the plasmid during cell division. Plasmids often carry multiple antibiotic resistance genes, contributing to the spread of multidrug-resistance (MDR). Antibiotic resistance mediated by MDR plasmids severely limits the treatment options for the infections caused by Gram-negative bacteria, especially family Enterobacteriaceae. The global spread of MDR plasmids has been enhanced by selective pressure from antimicrobial medications used in medical facilities and when raising animals for food.
Zygotic induction occurs when a bacterial cell carrying the silenced DNA of a bacterial virus in its chromosome transfers the viral DNA along with its own DNA to another bacterial cell lacking the virus, causing the recipient of the DNA to break open. In the donor cell, a repressor protein encoded by the prophage keeps the viral genes turned off so that virus is not produced. When DNA is transferred to the recipient cell by conjugation, the viral genes in the transferred DNA are immediately turned on because the recipient cell lacks the repressor. As a result, many virus are made in the recipient cell, and lysis eventually occurs to release the new virus.
Bacterial genetics is the subfield of genetics devoted to the study of bacterial genes. Bacterial genetics are subtly different from eukaryotic genetics, however bacteria still serve as a good model for animal genetic studies. One of the major distinctions between bacterial and eukaryotic genetics stems from the bacteria's lack of membrane-bound organelles, necessitating protein synthesis occur in the cytoplasm.
Integrative and conjugative elements (ICEs) are mobile genetic elements present in both gram-positive and gram-negative bacteria. In a donor cell, ICEs are located primarily on the chromosome, but have the ability to excise themselves from the genome and transfer to recipient cells via bacterial conjugation.