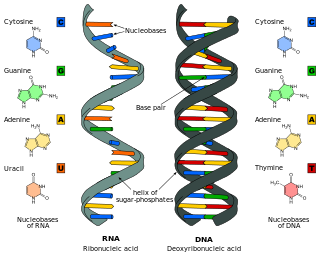
The RNA world is a hypothetical stage in the evolutionary history of life on Earth, in which self-replicating RNA molecules proliferated before the evolution of DNA and proteins. The term also refers to the hypothesis that posits the existence of this stage.
Virus classification is the process of naming viruses and placing them into a taxonomic system similar to the classification systems used for cellular organisms.
Hepatitis D is a type of viral hepatitis caused by the hepatitis delta virus (HDV). HDV is one of five known hepatitis viruses: A, B, C, D, and E. HDV is considered to be a satellite because it can propagate only in the presence of the hepatitis B virus (HBV). Transmission of HDV can occur either via simultaneous infection with HBV (coinfection) or superimposed on chronic hepatitis B or hepatitis B carrier state (superinfection).
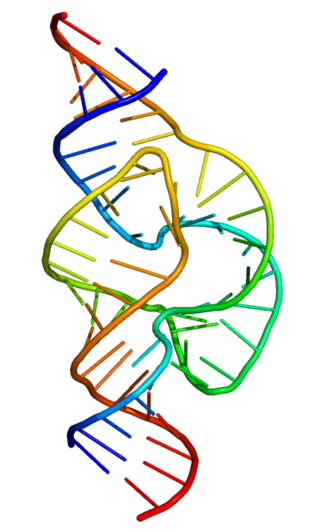
Ribozymes are RNA molecules that have the ability to catalyze specific biochemical reactions, including RNA splicing in gene expression, similar to the action of protein enzymes. The 1982 discovery of ribozymes demonstrated that RNA can be both genetic material and a biological catalyst, and contributed to the RNA world hypothesis, which suggests that RNA may have been important in the evolution of prebiotic self-replicating systems.
Virusoids are circular single-stranded RNA(s) dependent on viruses for replication and encapsidation. The genome of virusoids consists of several hundred (200–400) nucleotides and does not code for any proteins.
A satellite is a subviral agent that depends on the coinfection of a host cell with a helper virus for its replication. Satellites can be divided into two major classes: satellite viruses and satellite nucleic acids. Satellite viruses, which are most commonly associated with plants, are also found in mammals, arthropods, and bacteria. They encode structural proteins to enclose their genetic material, which are therefore distinct from the structural proteins of their helper viruses. Satellite nucleic acids, in contrast, do not encode their own structural proteins, but instead are encapsulated by proteins encoded by their helper viruses. The genomes of satellites range upward from 359 nucleotides in length for satellite tobacco ringspot virus RNA (STobRV).

Theodor Otto Diener was a Swiss-American plant pathologist. In 1971, he discovered that the causative agent of the potato spindle tuber disease is not a virus, but a novel agent, which consists solely of a short strand of single-stranded RNA without a protein capsid, eighty times smaller than the smallest viruses. He proposed to name it, and similar agents yet to be discovered, viroids. Viroids displaced viruses as the smallest known infectious agents.
The potato spindle tuber viroid (PSTVd) was the first viroid to be identified. PSTVd is a small, single stranded circular RNA molecule closely related to the chrysanthemum stunt viroid. Present within the viroidal RNA is the Pospiviroid RY motif stem loop common to its genus. The natural hosts are potatoes and tomatoes. All potatoes and tomatoes are susceptible to PSTVd and there is no form of natural resistance. Natural infections have also been seen in avocados and infections in other solanaceous crops have been induced in the laboratory. Until 2017 PSTVd was thought to be unable to infect Solanum sisymbriifolium. Then in May seeds exported by a Dutch company were noticed to be infected. These seeds were shipped from the company, but had been originally bred to their specifications in two Asian countries. Pstv also causes Tomato bunchy top and is seed transmitted in tomato.
The Pospiviroidae are a incertae sedis family of ssRNA viroids with 5 genera and 39 species, including the first viroid to be discovered, PSTVd, which is part of genus Pospiviroid. Their secondary structure is key to their biological activity. The classification of this family is based on differences in the conserved central region sequence. Pospiviroidae replication occurs in an asymmetric fashion via host cell RNA polymerase, RNase, and RNA ligase. Its hosts are plants, specifically dicotyledons and some monocotyledons.

Rolling circle replication (RCR) is a process of unidirectional nucleic acid replication that can rapidly synthesize multiple copies of circular molecules of DNA or RNA, such as plasmids, the genomes of bacteriophages, and the circular RNA genome of viroids. Some eukaryotic viruses also replicate their DNA or RNA via the rolling circle mechanism.

Potyvirus is a genus of positive-strand RNA viruses in the family Potyviridae. Plants serve as natural hosts. Like begomoviruses, members of this genus may cause significant losses in agricultural, pastoral, horticultural, and ornamental crops. More than 200 species of aphids spread potyviruses, and most are from the subfamily Aphidinae. The genus contains 190 species and potyviruses account for about thirty percent of all currently known plant viruses.
Non-cellular life, also known as acellular life, is life that exists without a cellular structure for at least part of its life cycle. Historically, most definitions of life postulated that an organism must be composed of one or more cells, but this is for some no longer considered necessary, and modern criteria allow for forms of life based on other structural arrangements.
Peach latent mosaic viroid is a species of the genus Pelamoviroid, which belongs to the family Avsunviroidae. This family is characterized as having chloroplastic viroids with hammerhead ribozymes. Peach latent mosaic viroid is a 336-351nt circular RNA which has a branched formation. This branched formation is stabilised by a pseudoknot between two kissing loops. Peach latent mosaic viroid was first described in the 1980s in Spain by a group of scientists.

The Avsunviroidae are a family of viroids. There are four species in three genera. They consist of RNA genomes between 246 and 375 nucleotides in length. They are single-stranded covalent circles and have intramolecular base pairing. All members lack a central conserved region.

The hammerhead ribozyme is an RNA motif that catalyzes reversible cleavage and ligation reactions at a specific site within an RNA molecule. It is one of several catalytic RNAs (ribozymes) known to occur in nature. It serves as a model system for research on the structure and properties of RNA, and is used for targeted RNA cleavage experiments, some with proposed therapeutic applications. Named for the resemblance of early secondary structure diagrams to a hammerhead shark, hammerhead ribozymes were originally discovered in two classes of plant virus-like RNAs: satellite RNAs and viroids. They are also known in some classes of retrotransposons, including the retrozymes. The hammerhead ribozyme motif has been ubiquitously reported in lineages across the tree of life.

The hairpin ribozyme is a small section of RNA that can act as a ribozyme. Like the hammerhead ribozyme it is found in RNA satellites of plant viruses. It was first identified in the minus strand of the tobacco ringspot virus (TRSV) satellite RNA where it catalyzes self-cleavage and joining (ligation) reactions to process the products of rolling circle virus replication into linear and circular satellite RNA molecules. The hairpin ribozyme is similar to the hammerhead ribozyme in that it does not require a metal ion for the reaction.

The hepatitis delta virus (HDV) ribozyme is a non-coding RNA found in the hepatitis delta virus that is necessary for viral replication and is the only known human virus that utilizes ribozyme activity to infect its host. The ribozyme acts to process the RNA transcripts to unit lengths in a self-cleavage reaction during replication of the hepatitis delta virus, which is thought to propagate by a double rolling circle mechanism. The ribozyme is active in vivo in the absence of any protein factors and was the fastest known naturally occurring self-cleaving RNA at the time of its discovery.
In virology, realm is the highest taxonomic rank established for viruses by the International Committee on Taxonomy of Viruses (ICTV), which oversees virus taxonomy. Six virus realms are recognized and united by specific highly conserved traits:

Ribozyviria is a realm of satellite nucleic acids — infectious agents that resemble viruses, but cannot replicate without a helper virus. Established in ICTV TaxoProp 2020.012D, the realm is named after the presence of genomic and antigenomic ribozymes of the Deltavirus type. The agents in Ribozyviria are satellite nucleic acids, which are distinct from satellite viruses in that they do not encode all of their own structural proteins but require proteins from their helper viruses in order to assemble. Additional common features include a rod-like structure, an RNA-binding "delta antigen" encoded in the genome, and animal hosts. Furthermore, the size range of the genomes of these viruses is between around 1547–1735nt, they encode a hammerhead ribozyme or a hepatitis delta virus ribozyme, and their coding capacity only involves one conserved protein. Most lineages of this realm are poorly understood, the notable exception being the genus Deltavirus, comprising the causal agents of hepatitis D in humans.
Retrozymes are a family of retrotransposons first discovered in the genomes of plants but now also known in genomes of animals. Retrozymes contain a hammerhead ribozyme (HHR) in their sequences, although they do not possess any coding regions. Retrozymes are nonautonomous retroelements, and so borrow proteins from other elements to move into new regions of a genome. Retrozymes are actively transcribed into covalently closed circular RNAs and are detected in both polarities, which may indicate the use of rolling circle replication in their lifecycle.