
An axon or nerve fiber is a long, slender projection of a nerve cell, or neuron, in vertebrates, that typically conducts electrical impulses known as action potentials away from the nerve cell body. The function of the axon is to transmit information to different neurons, muscles, and glands. In certain sensory neurons, such as those for touch and warmth, the axons are called afferent nerve fibers and the electrical impulse travels along these from the periphery to the cell body and from the cell body to the spinal cord along another branch of the same axon. Axon dysfunction can be the cause of many inherited and acquired neurological disorders that affect both the peripheral and central neurons. Nerve fibers are classed into three types – group A nerve fibers, group B nerve fibers, and group C nerve fibers. Groups A and B are myelinated, and group C are unmyelinated. These groups include both sensory fibers and motor fibers. Another classification groups only the sensory fibers as Type I, Type II, Type III, and Type IV.
In physiology, nociception, also nocioception; from Latin nocere 'to harm/hurt') is the sensory nervous system's process of encoding noxious stimuli. It deals with a series of events and processes required for an organism to receive a painful stimulus, convert it to a molecular signal, and recognize and characterize the signal to trigger an appropriate defensive response.

Itch is a sensation that causes a strong desire or reflex to scratch. Itches have resisted many attempts to be classified as any one type of sensory experience. Itches have many similarities to pain, and while both are unpleasant sensory experiences, their behavioral response patterns are different. Pain creates a withdrawal reflex, whereas itches leads to a scratch reflex.

In physiology, a stimulus is a detectable change in the physical or chemical structure of an organism's internal or external environment. The ability of an organism or organ to detect external stimuli, so that an appropriate reaction can be made, is called sensitivity (excitability). Sensory receptors can receive information from outside the body, as in touch receptors found in the skin or light receptors in the eye, as well as from inside the body, as in chemoreceptors and mechanoreceptors. When a stimulus is detected by a sensory receptor, it can elicit a reflex via stimulus transduction. An internal stimulus is often the first component of a homeostatic control system. External stimuli are capable of producing systemic responses throughout the body, as in the fight-or-flight response. In order for a stimulus to be detected with high probability, its level of strength must exceed the absolute threshold; if a signal does reach threshold, the information is transmitted to the central nervous system (CNS), where it is integrated and a decision on how to react is made. Although stimuli commonly cause the body to respond, it is the CNS that finally determines whether a signal causes a reaction or not.

A free nerve ending (FNE) or bare nerve ending, is an unspecialized, afferent nerve fiber sending its signal to a sensory neuron. Afferent in this case means bringing information from the body's periphery toward the brain. They function as cutaneous nociceptors and are essentially used by vertebrates to detect noxious stimuli that often result in pain.
A thermoreceptor is a non-specialised sense receptor, or more accurately the receptive portion of a sensory neuron, that codes absolute and relative changes in temperature, primarily within the innocuous range. In the mammalian peripheral nervous system, warmth receptors are thought to be unmyelinated C-fibres, while those responding to cold have both C-fibers and thinly myelinated A delta fibers. The adequate stimulus for a warm receptor is warming, which results in an increase in their action potential discharge rate. Cooling results in a decrease in warm receptor discharge rate. For cold receptors their firing rate increases during cooling and decreases during warming. Some cold receptors also respond with a brief action potential discharge to high temperatures, i.e. typically above 45 °C, and this is known as a paradoxical response to heat. The mechanism responsible for this behavior has not been determined.

A nociceptor is a sensory neuron that responds to damaging or potentially damaging stimuli by sending "possible threat" signals to the spinal cord and the brain. The brain creates the sensation of pain to direct attention to the body part, so the threat can be mitigated; this process is called nociception.

The spinothalamic tract is a part of the anterolateral system or the ventrolateral system, a sensory pathway to the thalamus. From the ventral posterolateral nucleus in the thalamus, sensory information is relayed upward to the somatosensory cortex of the postcentral gyrus.
Sensory neurons, also known as afferent neurons, are neurons in the nervous system, that convert a specific type of stimulus, via their receptors, into action potentials or graded receptor potentials. This process is called sensory transduction. The cell bodies of the sensory neurons are located in the dorsal ganglia of the spinal cord.

A dorsal root ganglion is a cluster of neurons in a dorsal root of a spinal nerve. The cell bodies of sensory neurons known as first-order neurons are located in the dorsal root ganglia.
Neuropathic pain is pain caused by a lesion or disease of the somatosensory nervous system. Neuropathic pain may be associated with abnormal sensations called dysesthesia or pain from normally non-painful stimuli (allodynia). It may have continuous and/or episodic (paroxysmal) components. The latter resemble stabbings or electric shocks. Common qualities include burning or coldness, "pins and needles" sensations, numbness and itching.
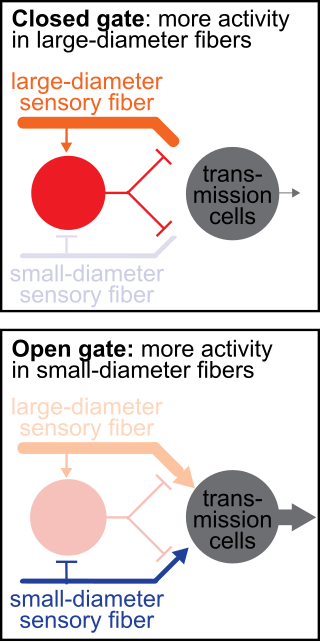
The gate control theory of pain asserts that non-painful input closes the nerve "gates" to painful input, which prevents pain sensation from traveling to the central nervous system.

Allodynia is a condition in which pain is caused by a stimulus that does not normally elicit pain. For example, sunburn can cause temporary allodynia, so that usually painless stimuli, such as wearing clothing or running cold or warm water over it, can be very painful. It is different from hyperalgesia, an exaggerated response from a normally painful stimulus. The term comes from Ancient Greek άλλος (állos) 'other', and οδύνη (odúnē) 'pain'.
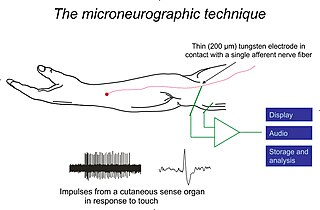
Microneurography is a neurophysiological method employed to visualize and record the traffic of nerve impulses that are conducted in peripheral nerves of waking human subjects. It can also be used in animal recordings. The method has been successfully employed to reveal functional properties of a number of neural systems, e.g. sensory systems related to touch, pain, and muscle sense as well as sympathetic activity controlling the constriction state of blood vessels. To study nerve impulses of an identified nerve, a fine tungsten needle microelectrode is inserted into the nerve and connected to a high input impedance differential amplifier. The exact position of the electrode tip within the nerve is then adjusted in minute steps until the electrode discriminates nerve impulses of interest. A unique feature and a significant strength of the microneurography method is that subjects are fully awake and able to cooperate in tests requiring mental attention, while impulses in a representative nerve fibre or set of nerve fibres are recorded, e.g. when cutaneous sense organs are stimulated or subjects perform voluntary precision movements.

Transient receptor potential cation channel subfamily M (melastatin) member 8 (TRPM8), also known as the cold and menthol receptor 1 (CMR1), is a protein that in humans is encoded by the TRPM8 gene. The TRPM8 channel is the primary molecular transducer of cold somatosensation in humans. In addition, mints can desensitize a region through the activation of TRPM8 receptors.

The axon reflex is the response stimulated by peripheral nerves of the body that travels away from the nerve cell body and branches to stimulate target organs. Reflexes are single reactions that respond to a stimulus making up the building blocks of the overall signaling in the body's nervous system. Neurons are the excitable cells that process and transmit these reflex signals through their axons, dendrites, and cell bodies. Axons directly facilitate intercellular communication projecting from the neuronal cell body to other neurons, local muscle tissue, glands and arterioles. In the axon reflex, signaling starts in the middle of the axon at the stimulation site and transmits signals directly to the effector organ skipping both an integration center and a chemical synapse present in the spinal cord reflex. The impulse is limited to a single bifurcated axon, or a neuron whose axon branches into two divisions and does not cause a general response to surrounding tissue.
Mechanosensation is the transduction of mechanical stimuli into neural signals. Mechanosensation provides the basis for the senses of light touch, hearing, proprioception, and pain. Mechanoreceptors found in the skin, called cutaneous mechanoreceptors, are responsible for the sense of touch. Tiny cells in the inner ear, called hair cells, are responsible for hearing and balance. States of neuropathic pain, such as hyperalgesia and allodynia, are also directly related to mechanosensation. A wide array of elements are involved in the process of mechanosensation, many of which are still not fully understood.
Zucapsaicin (Civanex) is a medication used to treat osteoarthritis of the knee and other neuropathic pain. Zucapsaicin is a member of phenols and a member of methoxybenzenes. It is a modulator of transient receptor potential cation channel subfamily V member 1 (TRPV-1), also known as the vanilloid or capsaicin receptor 1 that reduces pain, and improves articular functions. It is the cis-isomer of capsaicin. Civamide, manufactured by Winston Pharmaceuticals, is produced in formulations for oral, nasal, and topical use.
Group A nerve fibers are one of the three classes of nerve fiber as generally classified by Erlanger and Gasser. The other two classes are the group B nerve fibers, and the group C nerve fibers. Group A are heavily myelinated, group B are moderately myelinated, and group C are unmyelinated.

Presynaptic inhibition is a phenomenon in which an inhibitory neuron provides synaptic input to the axon of another neuron to make it less likely to fire an action potential. Presynaptic inhibition occurs when an inhibitory neurotransmitter, like GABA, acts on GABA receptors on the axon terminal. Or when endocannabinoids act as retrograde messengers by binding to presynaptic CB1 receptors, thereby indirectly modulating GABA and the excitability of dopamine neurons by reducing it and other presynaptic released neurotransmitters. Presynaptic inhibition is ubiquitous among sensory neurons.
