
The spirotrichs are a large and diverse group of ciliate protozoa. They typically have prominent oral cilia in the form of a series of polykinetids, called the adoral zone of membranelles, beginning anterior to the oral cavity and running down to the left side of the mouth. There may also be one or two paroral membranes on its right side. The body cilia are fused to form polykinetids called cirri in some, and are sparse to absent in others.

Paramecium is a genus of eukaryotic, unicellular ciliates, commonly studied as a model organism of the ciliate group. Paramecium are widespread in freshwater, brackish, and marine environments and are often abundant in stagnant basins and ponds. Because some species are readily cultivated and easily induced to conjugate and divide, they have been widely used in classrooms and laboratories to study biological processes. The usefulness of Paramecium as a model organism has caused one ciliate researcher to characterize it as the "white rat" of the phylum Ciliophora.

The Colpodea are a class of ciliates, of about 200 species common in freshwater and soil habitats. The body cilia are typically uniform, and are supported by dikinetids of characteristic structure, with cilia on both kinetosomes. The mouth may be apical or ventral, with more or less prominent associated polykinetids. Many are asymmetrical, the cells twisting sideways and then untwisting again prior to division, which often takes place within cysts. Colpoda, a kidney-shaped ciliate common in organic rich conditions, is representative.

The hypotrichs are a group of ciliated protozoa, common in fresh water, salt water, soil and moss. Hypotrichs possess compound ciliary organelles called "cirri," which are made up of thick tufts of cilia, sparsely distributed on the ventral surface of the cell. The multiple fused cilia which form a cirrus function together as a unit, enabling the organism to crawl along solid substrates such as submerged debris or sediments. Hypotrichs typically possess a large oral aperture, bordered on one side by a wreath or collar of membranelles, forming an "adoral zone of membranelles," or AZM.

Stylonychia is a genus of ciliates, in the subclass Hypotrichia. Species of Stylonychia are very common in fresh water and soil, and may be found on filamentous algae, surface films, and among particles of sediment. Like other Hypotrichs, Stylonychia has cilia grouped into membranelles alongside the mouth and cirri over the body. It is distinguished partly by long cirri at the posterior, usually a cluster of three. The largest can just be seen at a 25x magnification, and the smallest can just be seen at a 450x magnification.
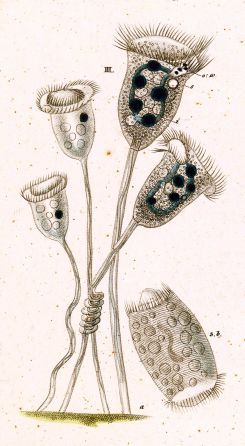
Vorticella is a genus of bell-shaped ciliates that have stalks to attach themselves to substrates. The stalks have contractile myonemes, allowing them to pull the cell body against substrates. The formation of the stalk happens after the free-swimming stage.

Spirostomum is a genus of ciliated protists in the class Heterotrichea. It is known for being very contractile. Having been first identified by Christian Gottfried Ehrenberg in 1834, further research has identified eight additional true morphospecies. This bacterivore genus mainly lives in the sediment deposits at the bottom of various aquatic habitats, and members possess rquA genes that could be responsible for their ability to survive in these hypoxic and anoxic environments. They are identifiable by their relatively large tubular/flat vermiform bodies. Their life cycle consists of a growth stage, in which they mature, and asexual and sexual reproduction stages. Some species are model organisms for studies on human pathogenic bacteria, while others are sensitive and accurate bioindicators for toxic substances.

Paramecium caudatum is a species of unicellular protist in the phylum Ciliophora. They can reach 0.33 mm in length and are covered with minute hair-like organelles called cilia. The cilia are used in locomotion and feeding. The species is very common, and widespread in marine, brackish and freshwater environments.
Karyorelictea is a class of ciliates in the subphylum Postciliodesmatophora. Most species are members of the microbenthos community, that is, microscopic organisms found in the marine interstitial habitat, though one genus, Loxodes, is found in freshwater.

The ciliates are a group of alveolates characterized by the presence of hair-like organelles called cilia, which are identical in structure to eukaryotic flagella, but are in general shorter and present in much larger numbers, with a different undulating pattern than flagella. Cilia occur in all members of the group and are variously used in swimming, crawling, attachment, feeding, and sensation.

Euplotes is a genus of ciliates in the subclass Euplotia. Species are widely distributed in marine and freshwater environments, as well as soil and moss. Most members of the genus are free-living, but two species have been recorded as commensal organisms in the digestive tracts of sea urchins.

Climacostomum is a genus of unicellular ciliates, belonging to the class Heterotrichea.

Pseudomonilicaryon anser is a species of unicellular ciliates in the family Dileptidae, also known under the names Dileptus anser and Dileptus cygnus. The species is common in fresh water ponds, stagnant pools, mosses and soils.
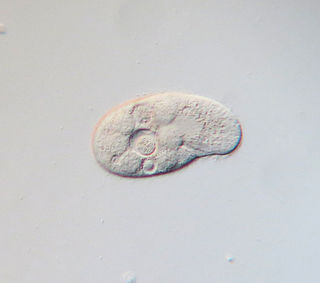
Colpidium colpoda are free-living ciliates commonly found in many freshwater environments including streams, rivers, lakes and ponds across the world. Colpidium colpoda is also frequently found inhabiting wastewater treatment plants. This species is used as an indicator of water quality and waste treatment plant performance.
Colpoda inflata is a unicellular organism, belonging to the genus Colpoda. Colpodeans are eucaryotic protozoans, that mainly feed on bacteria (bacteriophagous), vary a lot in size and have a funnel-shaped vestibule.

Condylostoma is a genus of unicellular ciliate protists, belonging to the class Heterotrichea.

Licnophora is a genus of ciliates in the family Licnophoridae. They typically have an hourglass-like shape and live as ectocommensals on marine animals.
Miamiensis avidus is a species of unicellular marine eukaryote that is a parasite of many different types of fish. It is one of several organisms known to cause the fish disease scuticociliatosis and is considered an economically significant pathogen of farmed fish. M. avidus is believed to be the cause of a 2017 die-off of fish and sharks in the San Francisco Bay.

Stentor roeselii is a free-living ciliate species of the genus Stentor, in the class Heterotrichea. It is a common and widespread protozoan, found throughout the world in freshwater ponds, lakes, rivers and ditches.
Parablepharismea is a class of free-living marine and brackish anaerobic ciliates that form a major clade of obligate anaerobes within the SAL group, together with the classes Muranotrichea and Armophorea.
