
Dinocephalosaurus is a genus of long necked, aquatic protorosaur that inhabited the Triassic seas of China. The genus contains the type and only known species, D. orientalis, which was named by Chun Li in 2003. Unlike other long-necked protorosaurs, Dinocephalosaurus convergently evolved a long neck not through elongation of individual neck vertebrae, but through the addition of neck vertebrae that each had a moderate length. As indicated by phylogenetic analyses, it belonged in a separate lineage that also included at least its closest relative Pectodens, which was named the Dinocephalosauridae in 2021. Like tanystropheids, however, Dinocephalosaurus probably used its long neck to hunt, utilizing the fang-like teeth of its jaws to ensnare prey; proposals that it employed suction feeding have not been universally accepted. It was probably a marine animal by necessity, as suggested by the poorly-ossified and paddle-like limbs which would have prevented it from going ashore.

Archosauromorpha is a clade of diapsid reptiles containing all reptiles more closely related to archosaurs rather than lepidosaurs. Archosauromorphs first appeared during the late Middle Permian or Late Permian, though they became much more common and diverse during the Triassic period.

Tanystropheus is an extinct genus of archosauromorph reptile which lived during the Triassic Period in Europe, Asia, and North America. It is recognisable by its extremely elongated neck, longer than the torso and tail combined. The neck was composed of 13 vertebrae strengthened by extensive cervical ribs. Tanystropheus is one of the most well-described non-archosauriform archosauromorphs, known from numerous fossils, including nearly complete skeletons. Some species within the genus may have reached a total length of 6 meters (20 ft), making Tanystropheus the longest non-archosauriform archosauromorph as well. Tanystropheus is the namesake of the family Tanystropheidae, a clade collecting many long-necked Triassic archosauromorphs previously described as "protorosaurs" or "prolacertiforms".
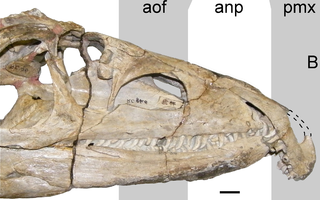
Proterosuchus is an extinct genus of archosauriform reptiles that lived during the Early Triassic. It contains three valid species: the type species P. fergusi and the referred species P. alexanderi and P. goweri. All three species lived in what is now South Africa. The genus was named in 1903 by the South African paleontologist Robert Broom. The genus Chasmatosaurus is a junior synonym of Proterosuchus.

Tasmaniosaurus is an extinct genus of archosauromorph reptile known from the Knocklofty Formation of West Hobart, Tasmania, Australia. The type species is T. triassicus. This genus is notable not only due to being one of the most complete Australian Triassic reptiles known, but also due to being a very close relative of Archosauriformes. Once believed to be a proterosuchid, this taxon is now believed to have been intermediate between advanced non-archosauriform archosauromorphs such as Prolacerta, and basal archosauriforms such as Proterosuchus. Features traditionally used to define Archosauria and later Archosauriformes, such as the presence of an antorbital fenestra and serrated teeth, are now known to have evolved prior to those groups due to their presence in Tasmaniosaurus.

Turfanosuchus is a genus of archosauriform reptile, likely a gracilisuchid archosaur, which lived during the Middle Triassic (Anisian) of northwestern China. The type species, T. dabanensis, was described by C.C. Young in 1973, based on a partially complete but disarticulated fossil skeleton found in the Kelamayi Formation of the Turfan Basin.

Vancleavea is a genus of extinct, armoured, non-archosaurian archosauriforms from the Late Triassic of western North America. The type and only known species is V. campi, named by Robert Long & Phillip A Murry in 1995. At that time, the genus was only known from fragmentary bones including osteoderms and vertebrae. However, since then many more fossils have been found, including a pair of nearly complete skeletons discovered in 2002. These finds have shown that members of the genus were bizarre semiaquatic reptiles. Vancleavea individuals had short snouts with large, fang-like teeth, and long bodies with small limbs. They were completely covered with bony plates known as osteoderms, which came in several different varieties distributed around the body. Phylogenetic analyses by professional paleontologists have shown that Vancleavea was an archosauriform, part of the lineage of reptiles that would lead to archosaurs such as dinosaurs and crocodilians. Vancleavea lacks certain traits which are present in most other archosauriforms, most notably the antorbital, mandibular and supratemporal fenestrae, which are weight-saving holes in the skulls of other taxa. However, other features clearly support its archosauriform identity, including a lack of intercentra, the presence of osteoderms, an ossified laterosphenoid, and several adaptations of the femur and ankle bones. In 2016, a new genus of archosauriform, Litorosuchus, was described. This genus resembled both Vancleavea and more typical archosauriforms in different respects, allowing Litorosuchus to act as a transitional fossil linking Vancleavea to less aberrant archosauriforms.

Rhadinosuchus is an extinct genus of proterochampsian archosauriform reptile from the Late Triassic. It is known only from the type species Rhadinosuchus gracilis, reposited in Munich, Germany. The fossil includes an incomplete skull and fragments of post-cranial material. Hosffstetter (1955), Kuhn (1966), Reig (1970) and Bonaparte (1971) hypothesized it to be synonymous with Cerritosaurus, but other characteristics suggest it is closer to Chanaresuchus and Gualosuchus, while it is certainly different from Proterochampsa and Barberenachampsa. The small size indicates it is a young animal, making it hard to classify.

Daemonosaurus is an extinct genus of possible theropod dinosaur from the Late Triassic of New Mexico. The only known fossil is a skull and neck fragments from deposits of the latest Triassic Chinle Formation at Ghost Ranch. Daemonosaurus was an unusual dinosaur with a short skull and large, fang-like teeth. It lived alongside early neotheropods such as Coelophysis, which would have been among the most common dinosaurs by the end of the Triassic. However, Daemonosaurus retains several plesiomorphic ("primitive") traits of the snout, and it likely lies outside the clade Neotheropoda. It may be considered a late-surviving basal theropod or non-theropod basal saurischian, possibly allied to other early predatory dinosaurs such as herrerasaurids or Tawa.

Jesairosaurus is an extinct genus of early archosauromorph reptile known from the Illizi Province of Algeria. It is known from a single species, Jesairosaurus lehmani. Although a potential relative of the long-necked tanystropheids, this lightly-built reptile could instead be characterized by its relatively short neck as well as various skull features.

Prolacerta is a genus of archosauromorph from the lower Triassic of South Africa and Antarctica. The only known species is Prolacerta broomi. The generic name Prolacerta is derived from Latin meaning “before lizard” and its species name broomi is in commemoration of the famous paleontologist Robert Broom, who discovered and studied many of the fossils found in rocks of the Karoo Supergroup. When first discovered, Prolacerta was considered to be ancestral to modern lizards, scientifically known as lacertilians. However, a study by Gow (1975) instead found that it shared more similarities with the lineage that would lead to archosaurs such as crocodilians and dinosaurs. Prolacerta is considered by modern paleontologists to be among the closest relatives of the Archosauriformes.

Palatodonta is an extinct genus of neodiapsid reptile known from the early Middle Triassic of the Netherlands. It was initially described in 2013 as a basal placodontiform closely related to a group of marine reptiles called placodonts, characterized by their crushing teeth and shell-like body armor. Under this interpretation, Palatodonta is transitional between placodonts and less specialized reptiles. Like placodonts, it has a row of large teeth on its palate, but while these teeth are thick and blunt in placodonts, Palatodonta has palatal teeth that are thin and pointed. A 2023 study instead classified it as a sauropterygomorph and the sister taxon to Eusaurosphargis. In other words, it is close to, but not within, Sauropterygia.

Pamelaria is an extinct genus of allokotosaurian archosauromorph reptile known from a single species, Pamelaria dolichotrachela, from the Middle Triassic of India. Pamelaria has sprawling legs, a long neck, and a pointed skull with nostrils positioned at the very tip of the snout. Among early archosauromorphs, Pamelaria is most similar to Prolacerta from the Early Triassic of South Africa and Antarctica. Both have been placed in the family Prolacertidae. Pamelaria, Prolacerta, and various other Permo-Triassic reptiles such as Protorosaurus and Tanystropheus have often been placed in a group of archosauromorphs called Protorosauria, which was regarded as one of the most basal group of archosauromorphs. However, more recent phylogenetic analyses indicate that Pamelaria and Prolacerta are more closely related to Archosauriformes than are Protorosaurus, Tanystropheus, and other protorosaurs, making Protorosauria a polyphyletic grouping.

Prolacertoides is an extinct genus of archosauromorph reptile from the Early Triassic of China, the type species being Prolacertoides jimusarensis. Prolacertoides means 'like Prolacerta', in reference to Prolacerta, another genus of archosauromorph which Prolacertoides was once believed to have been closely related to. Prolacertoides is known from a single partial skull, IVPP V3233, which was discovered in Xinjiang in northwestern China. The locality of its discovery belongs to the Cangfanggou Group of the Jiucaiyuan Formation, which is dated to the Induan age of the very early Triassic.

Asperoris is an extinct genus of archosauriform reptile known from the Middle Triassic Manda Beds of southwestern Tanzania. It is the first archosauriform known from the Manda Beds that is not an archosaur. However, its relationships with other non-archosaurian archosauriforms are uncertain. It was first named by Sterling J. Nesbitt, Richard J. Butler and David J. Gower in 2013 and the type species is Asperoris mnyama. Asperoris means "rough face" in Latin, referring to the distinctive rough texture of its skull bones.

Colobops is a genus of reptile from the Late Triassic of Connecticut. Only known from a tiny skull, this reptile has been interpreted to possess skull attachments for very strong jaw muscles. This may have given it a very strong bite, despite its small size. However, under some interpretations of the CT scan data, Colobops's bite force may not have been unusual compared to other reptiles. The generic name, Colobops, is a combination of κολοβός, meaning shortened, and ὤψ, meaning face. This translation, "shortened face", refers to its short and triangular skull. Colobops is known from a single species, Colobops noviportensis. The specific name, noviportensis, is a latinization of New Haven, the name of both the geological setting of its discovery as well as a nearby large city. The phylogenetic relations of Colobops are controversial. Its skull shares many features with those of the group Rhynchosauria, herbivorous archosauromorphs distantly related to crocodilians and dinosaurs. However, many of these features also resemble the skulls of the group Rhynchocephalia, an ancient order of reptiles including the modern tuatara, Sphenodon. Although rhynchosaurs and rhynchocephalians are not closely related and have many differences in the skeleton as a whole, their skulls are remarkably similar. As Colobops is only known from a skull, it is not certain which one of these groups it belonged to. Pritchard et al. (2018) interpreted it as a basal rhynchosaur, while Scheyer et al. (2020) reinterpreted it as a rhynchocephalian.
Boreopricea is an extinct genus of archosauromorph reptile from the Early Triassic of arctic Russia. It is known from a fairly complete skeleton discovered in a borehole on Kolguyev Island, though damage to the specimen and loss of certain bones has complicated study of the genus. Boreopricea shared many similarities with various other archosauromorphs, making its classification controversial. Various studies have considered it a close relative of Prolacerta, tanystropheids, both, or neither. Boreopricea is unique among early archosauromorphs due to possessing contact between the jugal and squamosal bones at the rear half of the skull.
Fraxinisaura is an extinct genus of basal lepidosauromorph reptile known from the Middle Triassic of Germany. The only known species is Fraxinisaura rozynekae. It possessed an elongated snout, unique features of the teeth, and an ilium which was intermediate in orientation between sphenodontians and squamates. Based on characteristics of the maxilla, it is considered a close relative of Marmoretta from the Middle Jurassic of the United Kingdom, resolving a ghost lineage between that genus and other Triassic basal lepidosauromorphs.

Rugarhynchos is an extinct genus of doswelliid archosauriform from the Late Triassic of New Mexico. The only known species is Rugarhynchos sixmilensis. It was originally described as a species of Doswellia in 2012, before receiving its own genus in 2020. Rugarhynchos was a close relative of Doswellia and shared several features with it, such as the absence of an infratemporal fenestra and heavily textured skull bones. However, it could also be distinguished by many unique characteristics, such as a thick diagonal ridge on the side of the snout, blunt spikes on its osteoderms, and a complex suture between the quadratojugal, squamosal, and jugal. Non-metric multidimensional scaling and tooth morphology suggest that Rugarhynchos had a general skull anatomy convergent with some crocodyliforms, spinosaurids, and phytosaurs. However, its snout was somewhat less elongated than those other reptiles.

Polymorphodon is an extinct genus of archosauriform reptile from the Middle Triassic of Germany. The only known species is Polymorphodon adorfi, discovered in Lower Keuper deposits at a quarry in Eschenau, Germany. Polymorphodon is notable for its heterodont dentition, with long and conical premaxillary teeth followed by thin maxillary teeth with large serrations. Maxillary teeth near the back of the mouth are short and leaf-shaped, similar to some living and extinct reptiles with a herbivorous or omnivorous diet. This may suggest that Polymorphodon had some reliance on plants in its diet, a rarity among basal archosauriforms, most of which are carnivores.



