
Dinocephalosaurus is a genus of long necked, aquatic protorosaur that inhabited the Triassic seas of China. The genus contains the type and only known species, D. orientalis, which was named by Chun Li in 2003. Unlike other long-necked protorosaurs, Dinocephalosaurus convergently evolved a long neck not through elongation of individual neck vertebrae, but through the addition of neck vertebrae that each had a moderate length. As indicated by phylogenetic analyses, it belonged in a separate lineage that also included at least its closest relative Pectodens, which was named the Dinocephalosauridae in 2021. Like tanystropheids, however, Dinocephalosaurus probably used its long neck to hunt, utilizing the fang-like teeth of its jaws to ensnare prey; proposals that it employed suction feeding have not been universally accepted. It was probably a marine animal by necessity, as suggested by the poorly-ossified and paddle-like limbs which would have prevented it from going ashore.

Archosauromorpha is a clade of diapsid reptiles containing all reptiles more closely related to archosaurs rather than lepidosaurs. Archosauromorphs first appeared during the late Middle Permian or Late Permian, though they became much more common and diverse during the Triassic period.

Tanystropheus is an extinct genus of archosauromorph reptile which lived during the Triassic Period in Europe, Asia, and North America. It is recognisable by its extremely elongated neck, longer than the torso and tail combined. The neck was composed of 13 vertebrae strengthened by extensive cervical ribs. Tanystropheus is one of the most well-described non-archosauriform archosauromorphs, known from numerous fossils, including nearly complete skeletons. Some species within the genus may have reached a total length of 6 meters (20 ft), making Tanystropheus the longest non-archosauriform archosauromorph as well. Tanystropheus is the namesake of the family Tanystropheidae, a clade collecting many long-necked Triassic archosauromorphs previously described as "protorosaurs" or "prolacertiforms".

Marasuchus is a genus of basal dinosauriform archosaur which is possibly synonymous with Lagosuchus. Both genera lived during the Late Triassic in what is now La Rioja Province, Argentina. Marasuchus contains a single species, Marasuchus lilloensis.

Erythrosuchidae are a family of large basal archosauriform carnivores that lived from the later Early Triassic (Olenekian) to the early Middle Triassic (Anisian).

Macrocnemus is an extinct genus of archosauromorph reptile known from the Middle Triassic of Europe and China. Macrocnemus is a member of the Tanystropheidae family and includes three species. Macrocnemus bassanii, the first species to be named and described, is known from the Besano Formation and adjacent paleontological sites in the Italian and Swiss Alps. Macrocnemus fuyuanensis, on the other hand, is known from the Zhuganpo Formation in southern China. A third species, Macrocnemus obristi, is known from the Prosanto Formation of Switzerland and is characterized by gracile limbs. The name Macrocnemus is Greek for "long tibia".

Turfanosuchus is a genus of archosauriform reptile, likely a gracilisuchid archosaur, which lived during the Middle Triassic (Anisian) of northwestern China. The type species, T. dabanensis, was described by C.C. Young in 1973, based on a partially complete but disarticulated fossil skeleton found in the Kelamayi Formation of the Turfan Basin.

The Sanga do Cabral Formation is an Early Triassic sedimentary rock formation found in Rio Grande do Sul, Brazil.
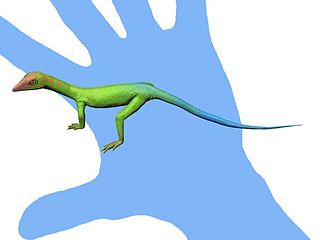
Cosesaurus is a genus of archosauromorph reptiles likely belonging to the family Tanystropheidae. It is known from fossil imprints of a single small skeleton, MGB V1, which was found in Muschelkalk outcrops near the municipalities of Mont-ral and Alcover in Spain. These outcrops are dated to the Ladinian age of the middle Triassic about 242 to 237 million years ago. The specimen is stored at the Museu Martorell, which is now part of the Museu de Ciències Naturals de Barcelona. The poor preservation and likely juvenile nature of the specimen has led to the anatomy of Cosesaurus being misidentified by several different sources. For example, Paul Ellenberger claimed that it was an ancestor to birds in the 1970s, while David Peters claimed that it was a pterosaur ancestor in 2000. Both of these claims contrast with mainstream scientific theories on the origins of either group, and other paleontologists who study the specimen are unable to find the features which Ellenberger or Peters reported to be present. The Ellenberger and Peters hypotheses are thus considered fringe theories with questionable scientific soundness due to their low reproducibility. Mainstream hypotheses for the relations of Cosesaurus generally agree that it is a "protorosaur", specifically a tanystropheid closely related to long-necked reptiles such as Macrocnemus, Tanytrachelos, Tanystropheus, or Langobardisaurus.

Jesairosaurus is an extinct genus of early archosauromorph reptile known from the Illizi Province of Algeria. It is known from a single species, Jesairosaurus lehmani. Although a potential relative of the long-necked tanystropheids, this lightly-built reptile could instead be characterized by its relatively short neck as well as various skull features.

Protorosauria is an extinct, likely paraphyletic group of basal archosauromorph reptiles from the latest Middle Permian to the end of the Late Triassic of Asia, Europe and North America. It was named by the English anatomist and paleontologist Thomas Henry Huxley in 1871 as an order, originally to solely contain Protorosaurus. Other names which were once considered equivalent to Protorosauria include Prolacertiformes and Prolacertilia.

Tanystropheidae is an extinct family of archosauromorph reptiles that lived throughout the Triassic Period, often considered to be "protorosaurs". They are characterized by their long, stiff necks formed from elongated cervical vertebrae with very long cervical ribs. Members of the group include both terrestrial and aquatic forms. While some tanystropheids were small lizard-like animals, other tanystropheids such as Tanystropheus were large animals that had necks that were several meters long, longer than the rest of their bodies.

Prolacertoides is an extinct genus of archosauromorph reptile from the Early Triassic of China, the type species being Prolacertoides jimusarensis. Prolacertoides means 'like Prolacerta', in reference to Prolacerta, another genus of archosauromorph which Prolacertoides was once believed to have been closely related to. Prolacertoides is known from a single partial skull, IVPP V3233, which was discovered in Xinjiang in northwestern China. The locality of its discovery belongs to the Cangfanggou Group of the Jiucaiyuan Formation, which is dated to the Induan age of the very early Triassic.
Nundasuchus is an extinct genus of crurotarsan, possibly a suchian archosaur related to Paracrocodylomorpha. Remains of this genus are known from the Middle Triassic Manda beds of southwestern Tanzania. It contains a single species, Nundasuchus songeaensis, known from a single partially complete skeleton, including vertebrae, limb elements, osteoderms, and skull fragments.
Aphanosauria is an extinct group of reptiles distantly related to dinosaurs. They are at the base of a group known as Avemetatarsalia, one of two main branches of archosaurs. The other main branch, Pseudosuchia, includes modern crocodilians. Aphanosaurs possessed features from both groups, indicating that they are the oldest and most primitive known clade of avemetatarsalians, at least in terms of their position on the archosaur family tree. Other avemetatarsalians include the flying pterosaurs, small bipedal lagerpetids, herbivorous silesaurids, and the incredibly diverse dinosaurs, which survive to the present day in the form of birds. Aphanosauria is formally defined as the most inclusive clade containing Teleocrater rhadinus and Yarasuchus deccanensis but not Passer domesticus or Crocodylus niloticus. This group was first recognized during the description of Teleocrater. Although only known by a few genera, Aphanosaurs had a widespread distribution across Pangaea in the Middle Triassic. They were fairly slow quadrupedal long-necked carnivores, a biology more similar to basal archosaurs than to advanced avemetatarsalians such as pterosaurs, lagerpetids, and early dinosaurs. In addition, they seemingly possess 'crocodile-normal' ankles, showing that 'advanced mesotarsal' ankles were not basal to the whole clade of Avemetatarsalia. Nevertheless, they possessed elevated growth rates compared to their contemporaries, indicating that they grew quickly, more like birds than other modern reptiles. Despite superficially resembling lizards, the closest modern relatives of aphanosaurs are birds.
Megacnemus is an extinct genus of archosauromorph reptile. Megacnemus is an enigmatic and poorly described genus, known from a single bone. The type species of Megacnemus, Megacnemus grandis, was named by Friedrich von Huene in 1954. The holotype of Megacnemus is a limb bone, believed to be a femur. Although the exact locality from which this bone was unearthed is unknown, it is believed to have come from Middle Triassic deposits near Gogolin, in southwest Poland. As the bone is more than 20 centimeters long, Megacnemus may have been quite large, similar to Vritraminosaurus in size.
Rhombopholis is an extinct genus of archosauromorph reptile known from England. The type species of Rhombopholis is Rhombopholis scutulata. Specimens of this genus were collected from the Leamington quarry, near Warwick. This locality belongs to the Bromsgrove Sandstone Formation, which is dated to the Anisian age of the Middle Triassic, approximately 245 million years ago.
Boreopricea is an extinct genus of archosauromorph reptile from the Early Triassic of arctic Russia. It is known from a fairly complete skeleton discovered in a borehole on Kolguyev Island, though damage to the specimen and loss of certain bones has complicated study of the genus. Boreopricea shared many similarities with various other archosauromorphs, making its classification controversial. Various studies have considered it a close relative of Prolacerta, tanystropheids, both, or neither. Boreopricea is unique among early archosauromorphs due to possessing contact between the jugal and squamosal bones at the rear half of the skull.
Trachelosauridae is an extinct clade of archosauromorph reptiles that lived throughout the Triassic period. Like their close relatives the tanystropheids, they were "protorosaur"-grade archosauromorphs characterized by their long necks. Unlike tanystropheids, which lengthen their neck primarily by elongating the individual cervical (neck) vertebrae, trachelosaurids achieved their long necks by the addition of more vertebrae. The most extreme example of this trend was Dinocephalosaurus, which had at least 32 vertebrae in the neck alone, far more than the 13 neck vertebrae of Tanystropheus.
Luxisaurus is an extinct genus of tanystropheid archosauromorph reptile from the Middle Triassic (Anisian) Guanling Formation of China. The genus contains a single species, L. terrestris, known from an articulated partial skeleton. Luxisaurus is hypothesized to have lived a more terrestrial lifestyle than many other tanystropheids, which may have been aquatic.






