A microswimmer is a microscopic object with the ability to move in a fluid environment.[1]Natural microswimmers are found everywhere in the natural world as biological microorganisms, such as bacteria, archaea, protists, sperm and microanimals. Since the turn of the millennium there has been increasing interest in manufacturing synthetic and biohybrid microswimmers. Although only two decades have passed since their emergence, they have already shown promise for various biomedical and environmental applications.[1]
Given the recent nature of the field, there is yet no consensus in the literature for the nomenclature of the microscopic objects this article refers to as "microswimmers". Among the many alternative names such objects are given in the literature, microswimmers, micro/nanorobots and micro/nanomotors are likely the most frequently encountered. Other common terms may be more descriptive, including information about the object shape, e.g., microtube or microhelix, its components, e.g., biohybrid, spermbot,[2] bacteriabot,[3] or micro-bio-robot,[4] or behavior, e.g., microrocket, microbullet, microtool or microroller. Researchers have also named their specific microswimmers e.g., medibots,[5] hairbots,[6] iMushbots,[7] IRONSperm,[8] teabots,[9] biobots,[10] T-budbots,[11] or MOFBOTS.[12][1]
Background
In 1828, the British biologist Robert Brown discovered the incessant jiggling motion of pollen in water and described his finding in his article "A Brief Account of Microscopical Observations…",[13] leading to extended scientific discussion about the origin of this motion. This enigma was resolved only in 1905, when Albert Einstein published his celebrated essay Über die von der molekularkinetischen Theorie der Wärme geforderte Bewegung von in ruhenden Flüssigkeiten suspendierten Teilchen.[14] Einstein not only deduced the diffusion of suspended particles in quiescent liquids, but also suggested these findings could be used to determine particle size — in a sense, he was the world's first microrheologist.[15]
Ever since Newton established his equations of motion, the mystery of motion on the microscale has emerged frequently in scientific history, as famously demonstrated by a couple of articles that should be discussed briefly. First, an essential concept, popularized by Osborne Reynolds, is that the relative importance of inertia and viscosity for the motion of a fluid depends on certain details of the system under consideration.[15] The Reynolds numberRe, named in his honor, quantifies this comparison as a dimensionless ratio of characteristic inertial and viscous forces:
E. M. Purcell
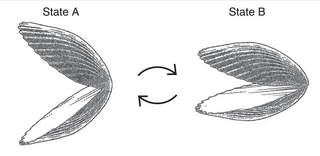
Purcell’s swimming scallop "Fast or slow, it exactly retraces its trajectory and it's back where it started".
Here, ρ represents the density of the fluid; u is a characteristic velocity of the system (for instance, the velocity of a swimming particle); l is a characteristic length scale (e.g., the swimmer size); and μ is the viscosity of the fluid. Taking the suspending fluid to be water, and using experimentally observed values for u, one can determine that inertia is important for macroscopic swimmers like fish (Re = 100), while viscosity dominates the motion of microscale swimmers like bacteria (Re = 10−4).[15]
The overwhelming importance of viscosity for swimming at the micrometer scale has profound implications for swimming strategy. This has been discussed memorably by E. M. Purcell, who invited the reader into the world of microorganisms and theoretically studied the conditions of their motion.[16] In the first place, propulsion strategies of large scale swimmers often involve imparting momentum to the surrounding fluid in periodic discrete events, such as vortex shedding, and coasting between these events through inertia. This cannot be effective for microscale swimmers like bacteria: due to the large viscous damping, the inertial coasting time of a micron-sized object is on the order of 1 μs. The coasting distance of a microorganism moving at a typical speed is about 0.1 angstroms (Å). Purcell concluded that only forces that are exerted in the present moment on a microscale body contribute to its propulsion, so a constant energy conversion method is essential.[16][15]
Microorganisms have optimized their metabolism for continuous energy production, while purely artificial microswimmers (microrobots) must obtain energy from the environment, since their on-board-storage-capacity is very limited. As a further consequence of the continuous dissipation of energy, biological and artificial microswimmers do not obey the laws of equilibrium statistical physics, and need to be described by non-equilibrium dynamics.[15] Mathematically, Purcell explored the implications of low Reynolds number by taking the Navier-Stokes equation and eliminating the inertial terms:
where is the velocity of the fluid and is the gradient of the pressure. As Purcell noted, the resulting equation — the Stokes equation — contains no explicit time dependence.[16] This has some important consequences for how a suspended body (e.g., a bacterium) can swim through periodic mechanical motions or deformations (e.g., of a flagellum). First, the rate of motion is practically irrelevant for the motion of the microswimmer and of the surrounding fluid: changing the rate of motion will change the scale of the velocities of the fluid and of the microswimmer, but it will not change the pattern of fluid flow. Secondly, reversing the direction of mechanical motion will simply reverse all velocities in the system. These properties of the Stokes equation severely restrict the range of feasible swimming strategies.[16][15]
As a concrete illustration, consider a mathematical scallop that consists of two rigid pieces connected by a hinge. Can the "scallop" swim by periodically opening and closing the hinge? No: regardless of how the cycle of opening and closing depends on time, the scallop will always return to its starting point at the end of the cycle. Here originated the striking quote: "Fast or slow, it exactly retraces its trajectory and it's back where it started".[16] In light of this scallop theorem, Purcell developed approaches concerning how artificial motion at the micro scale can be generated.[15] This paper continues to inspire ongoing scientific discussion; for example, recent work by the Fischer group from the Max Planck Institute for Intelligent Systems experimentally confirmed that the scallop principle is only valid for Newtonian fluids.[17][15]
Types
Different types of microswimmers are powered and actuated in different ways. Swimming strategies for individual microswimmers[3][18][19][20][21][22] as well as swarms of microswimmers[23][24][25][26][27][28] have been examined down through the years. Typically, microswimmers rely either on external power sources, as it is the case for magnetic,[29] optic,[10] or acoustic control,[30] or employ the fuel available in their surroundings, as is the case with biohybrid or catalytic microswimmers. Magnetic and acoustic actuation are typically compatible with in vivo microswimmer manipulation and catalytic microswimmers can be specifically engineered to employ in vivo fuels. The use of optical forces in biological fluids or in vivo is more challenging, but interesting examples have nevertheless been demonstrated.[10]
Often, researchers choose to take inspiration from nature, either for the entire microswimmer design, or for achieving a desired propulsion type. For example, one of the first bioinspired microswimmers consisted of human red blood cells modified with a flagellum-like artificial component made of filaments of magnetic particles bonded via biotin–streptavidin interactions.[31] More recently, biomimetic swimming inspired by worm-like travelling wave features,[32] shrimp locomotion,[33] and bacterial run-and-tumble motion,[34] was demonstrated by using shaped light.[10]
A different nature-inspired approach is the use of biohybrid microswimmers. These comprise a living component and a synthetic one. Biohybrids most often take advantage of the microscale motion of various biological systems and can also make use of other behaviours characterising the living component.[35] For magnetic bioinspired and biohybrid microswimmers, typical model organisms are bacteria, sperm cells and magnetotactic cells.[36] In addition to the use of magnetic forces, actuation of bioinspired microswimmers was also demonstrated using e.g., acoustic excitation[37] or optical forces.[38] Another nature-inspired behavior related to optical forces is that of phototaxis, which can be exploited by e.g., cargo-carrying microorganisms,[39] synthetic microswimmers[40][41][42] or biohybrid microswimmers.[43] A number of recent review papers are focused on explaining or comparing existing propulsion and control strategies used in microswimmer actuation.[44][45][46][47][48] Magnetic actuation is most often included for controlled in vivo guiding, even for microswimmers which rely on a different type of propulsion. In 2020, Koleoso et al. reviewed the use of magnetic small scale robots for biomedical applications and provide details about the various magnetic fields and actuation systems developed for such purposes.[29][1]
Strategies for the fabrication of microswimmers include two-photon polymerisation 3D printing, photolithography, template-assisted electrodeposition, or bonding of a living component to an inanimate one by exploiting different strategies. More recent approaches exploit 4D printing, which is the 3D printing of stimuli-responsive materials.[49][50][51][52] Further functionalization is often required, either to enable a certain type of actuation, e.g., metal coating for magnetic control or thermoplasmonic responses, or as part of the application, if certain characteristics are required for e.g., sensing, cargo transport, controlled interactions with the environment, or biodegradation.[53][54][55][56][1]
Some of the smallest known natural motile systems are motor proteins, i.e., proteins and protein complexes present in cells that carry out a variety of physiological functions by transducing chemical energy into mechanical energy. These motor proteins are classified as myosins, kinesins, or dyneins. Myosin motors are responsible for muscle contractions and the transport of cargousingactin filaments as tracks. Dynein motors and kinesin motors, on the other hand, use microtubules to transport vesicles across the cell.[59][60] The mechanism these protein motors use to convert chemical energy into movement depends on ATP hydrolysis, which leads to a conformation modification in the globular motor domain, leading to directed motion.[61][62][15]
Apart from motor proteins, enzymes, traditionally recognized for their catalytic functions in biochemical processes, can function as nanoscale machines that convert chemical energy into mechanical action at the molecular dimension. Diffusion of various enzymes (e.g. urease, and catalase), measured by fluorescent correlated spectroscopy (FCS), increases in a substrate-dependent manner.[63][64] Moreover, when enzymes are membrane-bound, their catalytic actions can drive lipid vesicle movement. For instance, lipid vesicles integrated with enzymes such as transmembrane adenosine 5’-triphosphatase, membrane-bound acid phosphatase, or urease exhibit enhanced mobility correlating with the enzymatic turnover rate.[65]
Bacteria can be roughly divided into two fundamentally different groups, gram-positive and gram-negative bacteria, distinguished by the architecture of their cell envelope. In each case the cell envelope is a complex multi-layered structure that protects the cell from its environment. In gram-positive bacteria, the cytoplasmic membrane is only surrounded by a thick cell wall of peptidoglycan. By contrast, the envelope of gram-negative bacteria is more complex and consists (from inside to outside) of the cytoplasmic membrane, a thin layer of peptidoglycan, and an additional outer membrane, also called the lipopolysaccharide layer. Other bacterial cell surface structures range from disorganised slime layers to highly structured capsules. These are made from secreted slimy or sticky polysaccharides or proteins that provide protection for the cells and are in direct contact with the environment. They have other functions, including attachment to solid surfaces. Additionally, protein appendages can be present on the surface: fimbriae and pili can have different lengths and diameters and their functions include adhesion and twitching motility.[66][67][15]
Specifically, for microorganisms that live in aqueous environments, locomotion refers to swimming, and hence the world is full of different classes of swimming microorganisms, such as bacteria, spermatozoa, protozoa, and algae. Bacteria move due to rotation of hair-like filaments called flagella, which are anchored to a protein motor complex on the bacteria cell wall.[15]
The following table, based on Schwarz et al., 2017,[68] lists some examples of natural or biological microswimmers.
"An artificial microswimmer is a cutting-edge technology with engineering and medical applications. A natural microswimmer, such as bacteria and sperm cells, also play important roles in wide varieties of engineering, medical and biological phenomena. Due to the small size of the microswimmer, the inertial effect of the surrounding flow field may be negligible. In such a case, reciprocal body deformation cannot induce migration of a swimmer, which is known as the scallop theorem. To overcome the implications of the scallop theorem, the microswimmer needs to undergo a nonreciprocal body deformation to achieve migration. The swimming strategy is thus completely different from macro-scale swimmers...".[90]
Under light fields, polystyrene/gold Janus particles are set to swim and rotate alternatively such that they follow a predefined path
One of the current engineering challenges is to create miniaturized functional vehicles that can carry out complex tasks at a small scale that would be otherwise impractical, inefficient, or outright impossible by conventional means. These vehicles are termed nano/micromotors or nano/microrobots, and should be distinguished from even smaller molecular machines for energy, computing, or other applications on the one side and static microelectromechanical systems (MEMS) on the other side of this size scale. Rather than being electronic devices on a chip, micromotors are able to move freely through a liquid medium while being steered or directed externally or by intrinsic design, which can be achieved by various mechanisms, most importantly catalytic reactions,[92][93][94][95]magnetic fields,[96] or ultrasonic waves.[97][98][99][100][101]
There are a variety of sensing, actuating, or pickup-and-delivery applications that scientists are currently aiming for, with local drug targeting for cancer treatment being one of the more prominent examples.[102][5] For applications like this, a micromotor needs to be able to move, i.e., to swim, freely in three dimensions efficiently controlled and directed with a reliable mechanism.[68]
It is a direct consequence of the small size scale of microswimmers that they have a low Reynolds number. This means the physics of how microswimmers swim is dominated by viscous drag forces, a problem which has been discussed extensively by physicists in the field.[99][103][58] This kind of swimming has challenged engineers as it is not commonly experienced in everyday life, but can nonetheless be observed in nature for motile microorganisms like sperm or certain bacteria. Naturally, these microorganisms served as inspiration from the very beginning to create artificial micromotors, as they were able to tackle the challenges that an active, self-sufficient microswimmer vehicle has to face.[104] With biomimetic approaches, researchers were able to imitate the flagella-based motion strategy of sperm and Escherichia coli bacteria by reproducing their respective flagellum shape and actuating it with magnetic fields.[31][105][68][15]
Microorganisms have adapted their locomotion to the harsh environment of low Reynolds number regime by invoking different swimming strategy.[106] For example, the E. coli moves by rotating its helical flagellum,[107][108]Chlamydomonas flagella have a breaststroke kind of motion.[109] African trypanosome has a helical flagellum attached to the cell body with a planar wave passing through it.[110][111] Swimming of these kind of natural swimmers have been investigated for the last half-century.[112] As a result of these studies, artificial swimmers have also been proposed, like Taylor sheet,[113] Purcell's two-hinge swimmer,[16][114] three-linked spheres swimmer,[115][116][117] elastic two-sphere swimmer[118] and three-sphere with a passive elastic arm,[119] which have further enhanced understanding about low Reynolds number swimmers. One of the challenges in proposing an artificial swimmer lies in the fact that the proposed movement stroke should not be reciprocal otherwise it cannot propel itself due to the Scallop theorem. In Scallop theorem, Purcell had argued that a swimmer with one-hinge or one degree of freedom is bound to perform reciprocal motion and thus will not be able to swim in the Stokes regime.[106][16][112]
Purcell proposed two possible ways to elude from Scallop theorem, one is 'corkscrew' motion[107][104] and the other is 'flexible oar' motion.[120][121] Using the concept of flexible oar, Dreyfus et al reported a micro swimmer that exploit elastic property of a slender filament made up of paramagnetic beads.[31] To break the time inversion symmetry, a passive head was attached to the flexible arm. The passive head reduces the velocity of the flexible swimmer, bigger the head, higher is the drag force experienced by the swimmer. The head is essential for swimming because without it the tail performs a reciprocal motion and the velocity of the swimmer reduces to zero.[122][112]
Another way microswimmers can propel is through catalytic reactions. Taking inspiration from Whitesides, who used the decomposition of hydrogen peroxide (H2O2) to propel cm/mm-scale objects on a water surface,[123] Sen et al. (2004) fabricated catalytic motors in the micrometer range.[92] These microswimmers were rod-shaped particles 370nm in diameter and consisted of 1μm long Pt and Au segments. They propelled via the decomposition of hydrogen peroxide in solution which would be catalyzed into water and oxygen. The Pt/Au rods were able to consistently reach speeds of up to 8μm/s in a solution of 3.3% hydrogen peroxide. The decomposition of hydrogen peroxide in the Pt side produces oxygen, two protons and two electrons. The two protons and electrons will travel towards the Au, where they will be used to react with another hydrogen peroxide molecule, to produce two water molecules. The movements of the two protons and the two electrons through the rod drag the fluid towards the Au side, thus this fluid flow will propel the rod in the opposite direction. This self-electrophoresis mechanism is what powers the motion of these rods.[93] Further analysis of the Pt/Au rods showed that they were capable of performing chemotaxis towards higher hydrogen peroxide concentrations,[94] transport cargo,[95] and exhibited steerable motion in an external magnetic field when inner Ni segments were added.[95]
Responding to stimuli
Symmetric self-thermophoretic active particle scale bar has a length of 1 μm
Reconfigurable synthetic or artificial microswimmers need internal feedback[125] Self-propelling microparticles are often proposed as synthetic models for biological microswimmers, yet they lack the internally regulated adaptation of their biological counterparts. Conversely, adaptation can be encoded in larger-scale soft-robotic devices but remains elusive to transfer to the colloidal scale.[125]
The ubiquity and success of motile bacteria are strongly coupled to their ability to autonomously adapt to different environments as they can reconfigure their shape, metabolism, and motility via internal feedback mechanisms.[126][127] Realizing artificial microswimmers with similar adaptation capabilities and autonomous behavior might substantially impact technologies ranging from optimal transport to sensing and microrobotics.[128] Focusing on adaptation, existing approaches at the colloidal scale mostly rely on external feedback, either to regulate motility via the spatiotemporal modulation of the propulsion velocity and direction[129][124][130][131] or to induce shape changes via the same magnetic or electric fields,[132][133][134] which are also driving the particles. On the contrary, endowing artificial microswimmers with an internal feedback mechanism, which regulates motility in response to stimuli that are decoupled from the source of propulsion, remains an elusive task.[125]
A promising route to achieve this goal is to exploit the coupling between particle shape and motility. Efficient switching between different propulsion states can, for instance, be reached by the spontaneous aggregation of symmetry-breaking active clusters of varying geometry,[135][136][137][138] albeit this process does not have the desired deterministic control. Conversely, designing colloidal clusters with fixed shapes and compositions offers fine control on motility[139][140][141] but lacks adaptation. Although progress on reconfigurable robots at the sub-millimeter scale has been made,[142][143][144][145][146] downscaling these concepts to the colloidal level demands alternative fabrication and design. Shape-shifting colloidal clusters reconfiguring along a predefined pathway in response to local stimuli[147] would combine both characteristics, with high potential toward the vision of realising adaptive artificial microswimmers.[125]
Bacterial biohybrid microswimmers development capture, delivery, sensing, and release
The so-called biohybrid microswimmer can be defined as a microswimmer that consist of both biological and artificial parts, for instance, one or several living microorganisms attached to one or various synthetic parts. The biohybrid approach directly employs living microorganisms to be a main component or modified base of a functional microswimmer.[150][151] Initially microorganisms were used as the motor units for artificial devices, but in recent years this role has been extended and modified toward other functionalities that take advantage of the biological capabilities of these organisms considering their means of interacting with other cells and living matter, specifically for applications inside the human body like drug delivery or fertilisation.[152][153][68]
A distinct advantage of microorganisms is that they naturally integrate motility and various biological functions in a conveniently miniaturised package, coupled with autonomous sensing and decision-making capabilities. They are able to adapt and thrive in complex in vivo environments and are capable of self-repair and self-assembly upon interaction with their surroundings. In that sense, self-sufficient microorganisms naturally function very similar to what we envision for artificially created microrobots: They harvest chemical energy from their surroundings to power molecular motor proteins that serve as actuators, they employ ion channels and microtubular networks to act as intracellular wiring, they rely on RNA or DNA as memory for control algorithms, and they feature an array of various membrane proteins to sense and evaluate their surroundings. All these abilities act together to allow microbes to thrive and pursue their goal and function. In principle, these abilities also qualify them as biological microrobots for novel operations like theranostics, the combination of diagnosis and therapy, if we are able to impose such functions artificially, for example, by functionalisation with therapeutics. Further, artificial extensions may be used as handles for external control and supervision mechanisms or to enhance the microbe's performance to guide and tailor its functions for specific applications.[68]
In fact, the biohybrid approach can be conceived in a dualistic way, with respect to the three basic ingredients of an in vivo microrobot, which are motility, control, and functionality. Figure 1 illustrates how these three ingredients can be either realized biologically, i.e., by the microorganism, or artificially, i.e., by the synthetic component. For example, a hybrid biomicromotor based on a sperm cell can be driven by the flagellum of the sperm or by an attached artificial helical flagellum.[154][155] It can orient itself autonomously via biological interactions with its surroundings and other cells, or be controlled and supervised externally via artificial sensors and actuators. Finally, it can carry out a biological function, like its inherent ability to fertilize an egg cell, or an artificially imposed function, like the delivery of synthetic drugs or DNA vectors. A biohybrid device may deploy any feasible combination of such biological and artificial components in order to carry out a specific application.[68]
Navigation
Hydrodynamics can determine the optimal route for microswimmer navigation[156] Compared to the well explored problem of how to steer a macroscopic agent, like an airplane or a moon lander, to optimally reach a target, optimal navigation strategies for microswimmers experiencing hydrodynamic interactions with walls and obstacles are far-less understood.[156] Furthermore, hydrodynamic interactions in suspensions of microswimmers produce complex behavior.[157][158] The quest on how to navigate or steer to optimally reach a target is important, e.g., for airplanes to save fuel while facing complex wind patterns on their way to a remote destination, or for the coordination of the motion of the parts of a space-agent to safely land on the moon. These classical problems are well-explored and are usually solved using optimal control theory.[159] Likewise, navigation and search strategies are frequently encountered in a plethora of biological systems, including the foraging of animals for food,[160] or of T cells searching for targets to mount an immune response.[161]
There is growing interest in optimal navigation problems and search strategies[162][163][164][165][166][167] of microswimmers[58][103][168][169] and "dry" active Brownian particles,[170][99][171][172][156] The general problem regarding the optimal trajectory of a microswimmer which can freely steer but cannot control its speed toward a predefined target (point-to-point navigation) can be referred to as "the optimal microswimmer navigation problem". The characteristic differences between the optimal microswimmer navigation problem and conventional optimal control problems for macroagents like airplanes, cruise-ships, or moon-landers root in the presence of a low-Reynolds-number solvent in the former problem only. They comprise (i) overdamped dynamics, (ii) thermal fluctuations, and (iii) long-ranged fluid-mediated hydrodynamic interactions with interfaces, walls, and obstacles, all of which are characteristic for microswimmers.[99] In particular, the non-conservative hydrodynamic forces which microswimmers experience call for a distinct navigation strategy than the conservative gravitational forces acting, e.g. on space vehicles. Recent work has explored optimal navigation problems of dry active particles (and particles in external flow fields) accounting for (i) and partly also for (ii). Specifically recent research has pioneered the use of reinforcement learning[173][174][175] such as determining optimal steering strategies of active particles to optimally navigate toward a target position[162][163][166][167] or to exploit external flow fields to avoid getting trapped in certain flow structures by learning smart gravitaxis.[176] Deep reinforcement learning has been used to explore microswimmer navigation problems in mazes and obstacle arrays[177] assuming global[163] or only local[164] knowledge of the environment. Analytical approaches to optimal active particle navigation[165][166] complement these works and allow testing machine-learned results.[166][167][156]
Applications
As is the case for microtechnology and nanotechnology in general, the history of microswimmer applications arguably starts with Richard Feynman’s famous lecture There's Plenty of Room at the Bottom.[178] In the visionary speech, among other topics, Feynman addressed the idea of microscopic surgeons, saying: "...it would be interesting in surgery if you could swallow the surgeon. You put the mechanical surgeon inside the blood vessel and it goes into the heart and <<looks>> around (of course the information has to be fed out). It finds out which valve is the faulty one and takes a little knife and slices it out. Other small machines might be permanently incorporated in the body to assist some inadequately-functioning organ." The concept of the surgeon one could swallow was soon after presented in the science-fiction movie Fantastic Voyage and in Isaac Asimov’s writings.[1]
Only a few decades later, microswimmers aiming to become true microscale surgeons evolved from an intriguing science-fiction concept to a reality explored in many research laboratories around the world, as already highlighted by Metin Sitti in 2009.[180][1] These active agents that can self-propel in a low Reynolds number environment might play a key role in the future of nanomedicine, as popularised in 2016 by Yuval Noah Harari in Homo Deus: A Brief History of Tomorrow.[181] In particular, they might become useful for the targeted delivery of genes[182] or drugs[183][184] and other cargo[185][186] to a certain target (e.g. a cancer cell) through our blood vessels, requiring them to find a good, or ideally optimal, path toward the target avoiding, e.g., obstacles and unfortunate flow field regions.[156]
Already in 2010, Nelson et al. reviewed the existing and envisioned applications of microrobots in minimally invasive medicine.[187] Since then, the field has grown, and it has become clear that microswimmers have much potential for biomedical applications.[1] Already, many interesting tasks can be performed in vitro using tailored microswimmers. Still, as of 2020, a number of challenges regarding in vivo control, biocompatibility and long-term biosafety need to be overcome before microswimmers can become a viable option for many clinical applications.[188][1]
A schematic representation of the classification of biomedical applications is shown in the diagram on the left below. This includes the use of microswimmers for cargo transport in drug delivery and other biomedical applications, as well as assisted fertilisation, sensing, micromanipulation and imaging. Some of the more complex microswimmers fit into multiple categories, as they are applied simultaneously for e.g., sensing and drug delivery.[1]
Biomedical applications of microswimmers
Essentials for a microswimmer to function with medical interventional capabilities
The design of an untethered microscopic mobile machine or microrobot to function in vivo with medical interventional capabilities should assume an integrated approach where design 3D body shape, material composition, manufacturing technique, deployment strategy, actuation and control methods, imaging modality, permeation of biological barriers, and the execution of the prescribed medical tasks need to be considered altogether, as illustrated in the diagram on the right above. Each of these essential aspects contains a special design consideration, which must be reflected at the physical design of the microrobot.[189]
Microfluidics refers to a system that manipulates a small amount of fluids using small channels with sizes ten to hundreds micrometres. It is a multidisciplinary field that involves molecular analysis, molecular biology, and microelectronics. It has practical applications in the design of systems that process low volumes of fluids to achieve multiplexing, automation, and high-throughput screening. Microfluidics emerged in the beginning of the 1980s and is used in the development of inkjet printheads, DNA chips, lab-on-a-chip technology, micro-propulsion, and micro-thermal technologies.
Microbotics is the field of miniature robotics, in particular mobile robots with characteristic dimensions less than 1 mm. The term can also be used for robots capable of handling micrometer size components.
Nanoid robotics, or for short, nanorobotics or nanobotics, is an emerging technology field creating machines or robots, which are called nanorobots or simply nanobots, whose components are at or near the scale of a nanometer. More specifically, nanorobotics refers to the nanotechnology engineering discipline of designing and building nanorobots with devices ranging in size from 0.1 to 10 micrometres and constructed of nanoscale or molecular components. The terms nanobot, nanoid, nanite, nanomachine and nanomite have also been used to describe such devices currently under research and development.
Motility is the ability of an organism to move independently, using metabolic energy.
A nanomotor is a molecular or nanoscale device capable of converting energy into movement. It can typically generate forces on the order of piconewtons.
Molecular motors are natural (biological) or artificial molecular machines that are the essential agents of movement in living organisms. In general terms, a motor is a device that consumes energy in one form and converts it into motion or mechanical work; for example, many protein-based molecular motors harness the chemical free energy released by the hydrolysis of ATP in order to perform mechanical work. In terms of energetic efficiency, this type of motor can be superior to currently available man-made motors. One important difference between molecular motors and macroscopic motors is that molecular motors operate in the thermal bath, an environment in which the fluctuations due to thermal noise are significant.
Magnetic nanoparticles (MNPs) are a class of nanoparticle that can be manipulated using magnetic fields. Such particles commonly consist of two components, a magnetic material, often iron, nickel and cobalt, and a chemical component that has functionality. While nanoparticles are smaller than 1 micrometer in diameter, the larger microbeads are 0.5–500 micrometer in diameter. Magnetic nanoparticle clusters that are composed of a number of individual magnetic nanoparticles are known as magnetic nanobeads with a diameter of 50–200 nanometers. Magnetic nanoparticle clusters are a basis for their further magnetic assembly into magnetic nanochains. The magnetic nanoparticles have been the focus of much research recently because they possess attractive properties which could see potential use in catalysis including nanomaterial-based catalysts, biomedicine and tissue specific targeting, magnetically tunable colloidal photonic crystals, microfluidics, magnetic resonance imaging, magnetic particle imaging, data storage, environmental remediation, nanofluids, optical filters, defect sensor, magnetic cooling and cation sensors.
Bacterial motility is the ability of bacteria to move independently using metabolic energy. Most motility mechanisms that evolved among bacteria also evolved in parallel among the archaea. Most rod-shaped bacteria can move using their own power, which allows colonization of new environments and discovery of new resources for survival. Bacterial movement depends not only on the characteristics of the medium, but also on the use of different appendages to propel. Swarming and swimming movements are both powered by rotating flagella. Whereas swarming is a multicellular 2D movement over a surface and requires the presence of surfactants, swimming is movement of individual cells in liquid environments.
Active matter is matter composed of large numbers of active "agents", each of which consumes energy in order to move or to exert mechanical forces. Such systems are intrinsically out of thermal equilibrium. Unlike thermal systems relaxing towards equilibrium and systems with boundary conditions imposing steady currents, active matter systems break time reversal symmetry because energy is being continually dissipated by the individual constituents. Most examples of active matter are biological in origin and span all the scales of the living, from bacteria and self-organising bio-polymers such as microtubules and actin, to schools of fish and flocks of birds. However, a great deal of current experimental work is devoted to synthetic systems such as artificial self-propelled particles. Active matter is a relatively new material classification in soft matter: the most extensively studied model, the Vicsek model, dates from 1995.
In physics, the scallop theorem states that a swimmer that performs a reciprocal motion cannot achieve net displacement in a low-Reynolds number Newtonian fluid environment, i.e. a fluid that is highly viscous. Such a swimmer deforms its body into a particular shape through a sequence of motions and then reverts to the original shape by going through the sequence in reverse. At low Reynolds number, time or inertia does not come into play, and the swimming motion is purely determined by the sequence of shapes that the swimmer assumes.
Self-propelled particles (SPP), also referred to as self-driven particles, are terms used by physicists to describe autonomous agents, which convert energy from the environment into directed or persistent motion. Natural systems which have inspired the study and design of these particles include walking, swimming or flying animals. Other biological systems include bacteria, cells, algae and other micro-organisms. Generally, self-propelled particles often refer to artificial systems such as robots or specifically designed particles such as swimming Janus colloids, bimetallic nanorods, nanomotors and walking grains. In the case of directed propulsion, which is driven by a chemical gradient, this is referred to as chemotaxis, observed in biological systems, e.g. bacteria quorum sensing and ant pheromone detection, and in synthetic systems, e.g. enzyme molecule chemotaxis and enzyme powered hard and soft particles.
Robotic sperm are biohybrid microrobots consisting of sperm cells and artificial microstructures. Currently there are two types of spermbots. The first type, the tubular spermbot, consists of a single sperm cell that is captured inside a microtube. Single bull sperm cells enter these microtubes and become trapped inside. The tail of the sperm is the driving force for the microtube. The second type, the helical spermbot, is a small helix structure which captures and transports single immotile sperm cells. In this case, a rotating magnetic field drives the helix in a screw-like motion. Both kinds of spermbots can be guided by weak magnetic fields. These two spermbot designs are hybrid microdevices, they consist of a living cell combined with synthetic attachments. Other approaches exist to create purely synthetic microdevices inspired by the swimming of natural sperm cells, i.e. with a biomimetic design, for example so-called Magnetosperm which are made of a flexible polymeric structure coated with a magnetic layer and can be actuated by a magnetic field.
Many experimental realizations of self-propelled particles exhibit a strong tendency to aggregate and form clusters, whose dynamics are much richer than those of passive colloids. These aggregates of particles form for a variety of reasons, from chemical gradients to magnetic and ultrasonic fields. Self-propelled enzyme motors and synthetic nanomotors also exhibit clustering effects in the form of chemotaxis. Chemotaxis is a form of collective motion of biological or non-biological particles toward a fuel source or away from a threat, as observed experimentally in enzyme diffusion and also synthetic chemotaxis or phototaxis. In addition to irreversible schooling, self-propelled particles also display reversible collective motion, such as predator–prey behavior and oscillatory clustering and dispersion.
Collective motion is defined as the spontaneous emergence of ordered movement in a system consisting of many self-propelled agents. It can be observed in everyday life, for example in flocks of birds, schools of fish, herds of animals and also in crowds and car traffic. It also appears at the microscopic level: in colonies of bacteria, motility assays and artificial self-propelled particles. The scientific community is trying to understand the universality of this phenomenon. In particular it is intensively investigated in statistical physics and in the field of active matter. Experiments on animals, biological and synthesized self-propelled particles, simulations and theories are conducted in parallel to study these phenomena. One of the most famous models that describes such behavior is the Vicsek model introduced by Tamás Vicsek et al. in 1995.
Bradley James Nelson is an American roboticist and entrepreneur. He has been the Professor of Robotics and Intelligent Systems at ETH Zurich since 2002 and is known for his research in microrobotics, nanorobotics, and medical robotics.
Self-propulsion is the autonomous displacement of nano-, micro- and macroscopic natural and artificial objects, containing their own means of motion. Self-propulsion is driven mainly by interfacial phenomena. Various mechanisms of self-propelling have been introduced and investigated, which exploited phoretic effects, gradient surfaces, breaking the wetting symmetry of a droplet on a surface, the Leidenfrost effect, the self-generated hydrodynamic and chemical fields originating from the geometrical confinements, and soluto- and thermo-capillary Marangoni flows. Self-propelled system demonstrate a potential as micro-fluidics devices and micro-mixers. Self-propelled liquid marbles have been demonstrated.
Marine prokaryotes are marine bacteria and marine archaea. They are defined by their habitat as prokaryotes that live in marine environments, that is, in the saltwater of seas or oceans or the brackish water of coastal estuaries. All cellular life forms can be divided into prokaryotes and eukaryotes. Eukaryotes are organisms whose cells have a nucleus enclosed within membranes, whereas prokaryotes are the organisms that do not have a nucleus enclosed within a membrane. The three-domain system of classifying life adds another division: the prokaryotes are divided into two domains of life, the microscopic bacteria and the microscopic archaea, while everything else, the eukaryotes, become the third domain.
Protists are the eukaryotes that cannot be classified as plants, fungi or animals. They are mostly unicellular and microscopic. Many unicellular protists, particularly protozoans, are motile and can generate movement using flagella, cilia or pseudopods. Cells which use flagella for movement are usually referred to as flagellates, cells which use cilia are usually referred to as ciliates, and cells which use pseudopods are usually referred to as amoeba or amoeboids. Other protists are not motile, and consequently have no built-in movement mechanism.
A biohybrid microswimmer also known as biohybrid nanorobot, can be defined as a microswimmer that consist of both biological and artificial constituents, for instance, one or several living microorganisms attached to one or various synthetic parts.
Run-and-tumble motion is a movement pattern exhibited by certain bacteria and other microscopic agents. It consists of an alternating sequence of "runs" and "tumbles": during a run, the agent propels itself in a fixed direction, and during a tumble, it remains stationary while it reorients itself in preparation for the next run.
↑ Brown, James F. (1852). "XXIV. On some salts and products of decomposition of pyromeconic acid". The London, Edinburgh, and Dublin Philosophical Magazine and Journal of Science. 4 (24): 161–168. doi:10.1080/14786445208647098.
↑ Abbott, Jake J.; Peyer, Kathrin E.; Lagomarsino, Marco Cosentino; Zhang, Li; Dong, Lixin; Kaliakatsos, Ioannis K.; Nelson, Bradley J. (2009). "How Should Microrobots Swim?". The International Journal of Robotics Research. 28 (11–12): 1434–1447. doi:10.1177/0278364909341658. S2CID62330062.
↑ Schamel, Debora; Mark, Andrew G.; Gibbs, John G.; Miksch, Cornelia; Morozov, Konstantin I.; Leshansky, Alexander M.; Fischer, Peer (2014). "Nanopropellers and Their Actuation in Complex Viscoelastic Media". ACS Nano. 8 (9): 8794–8801. doi:10.1021/nn502360t. PMID24911046.
↑ Rao, K. Jagajjanani; Li, Fei; Meng, Long; Zheng, Hairong; Cai, Feiyan; Wang, Wei (2015). "A Force to be Reckoned with: A Review of Synthetic Microswimmers Powered by Ultrasound". Small. 11 (24): 2836–2846. doi:10.1002/smll.201403621. PMID25851515.
↑ Luo, Ming; Feng, Youzeng; Wang, Tingwei; Guan, Jianguo (2018). "Micro-/Nanorobots at Work in Active Drug Delivery". Advanced Functional Materials. 28 (25). doi:10.1002/adfm.201706100. S2CID104145610.
↑ Vogel, Pia D. (2005). "Nature's design of nanomotors". European Journal of Pharmaceutics and Biopharmaceutics. 60 (2): 267–277. doi:10.1016/j.ejpb.2004.10.007. PMID15939237.
↑ Kim, Dal Hyung; Cheang, U. Kei; Kőhidai, László; Byun, Doyoung; Kim, Min Jun (2010). "Artificial magnetotactic motion control of Tetrahymena pyriformis using ferromagnetic nanoparticles: A tool for fabrication of microbiorobots". Applied Physics Letters. 97 (17): 173702. Bibcode:2010ApPhL..97q3702K. doi:10.1063/1.3497275.
1 2 Maree, L.; Van Der Horst, G. (2013). "Quantification and identification of sperm subpopulations using computer-aided sperm analysis and species-specific cut-off values for swimming speed". Biotechnic & Histochemistry. 88 (3–4): 181–193. doi:10.3109/10520295.2012.757366. hdl:10566/3120. PMID23331185. S2CID19603301.
↑ Yang, Yuguang; Bevan, Michael A.; Li, Bo (2020). "Micro/Nano Motor Navigation and Localization via Deep Reinforcement Learning". Advanced Theory and Simulations. 3 (6). arXiv:2002.06775. doi:10.1002/adts.202000034. S2CID211133324.
↑ Feynman, R. (2018). "There’s plenty of room at the bottom". In: Hey, Anthony (2018). Feynman and computation: exploring the limits of computers. Boca Raton: CRC Press. pp.63–76. ISBN978-0-429-50045-9.
↑ Demirörs, Ahmet F.; Akan, Mehmet Tolga; Poloni, Erik; Studart, André R. (2018). "Active cargo transport with Janus colloidal shuttles using electric and magnetic fields". Soft Matter. 14 (23): 4741–4749. Bibcode:2018SMat...14.4741D. doi:10.1039/C8SM00513C. PMID29799053.
↑ Nelson, Bradley J.; Kaliakatsos, Ioannis K.; Abbott, Jake J. (2010). "Microrobots for Minimally Invasive Medicine". Annual Review of Biomedical Engineering. 12: 55–85. doi:10.1146/annurev-bioeng-010510-103409. PMID20415589.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.