
Mitosis is a part of the cell cycle in which replicated chromosomes are separated into two new nuclei. Cell division by mitosis is an equational division which gives rise to genetically identical cells in which the total number of chromosomes is maintained. Mitosis is preceded by the S phase of interphase and is followed by telophase and cytokinesis; which divides the cytoplasm, organelles and cell membrane of one cell into two new cells containing roughly equal shares of these cellular components. The different stages of mitosis altogether define the mitotic phase of a cell cycle—the division of the mother cell into two daughter cells genetically identical to each other.

Microtubules are polymers of tubulin that form part of the cytoskeleton and provide structure and shape to eukaryotic cells. Microtubules can be as long as 50 micrometres, as wide as 23 to 27 nm and have an inner diameter between 11 and 15 nm. They are formed by the polymerization of a dimer of two globular proteins, alpha and beta tubulin into protofilaments that can then associate laterally to form a hollow tube, the microtubule. The most common form of a microtubule consists of 13 protofilaments in the tubular arrangement.

Cytokinesis is the part of the cell division process during which the cytoplasm of a single eukaryotic cell divides into two daughter cells. Cytoplasmic division begins during or after the late stages of nuclear division in mitosis and meiosis. During cytokinesis the spindle apparatus partitions and transports duplicated chromatids into the cytoplasm of the separating daughter cells. It thereby ensures that chromosome number and complement are maintained from one generation to the next and that, except in special cases, the daughter cells will be functional copies of the parent cell. After the completion of the telophase and cytokinesis, each daughter cell enters the interphase of the cell cycle.

In cell biology, the spindle apparatus is the cytoskeletal structure of eukaryotic cells that forms during cell division to separate sister chromatids between daughter cells. It is referred to as the mitotic spindle during mitosis, a process that produces genetically identical daughter cells, or the meiotic spindle during meiosis, a process that produces gametes with half the number of chromosomes of the parent cell.

Clathrin is a protein that plays a major role in the formation of coated vesicles. Clathrin was first isolated by Barbara Pearse in 1976. It forms a triskelion shape composed of three clathrin heavy chains and three light chains. When the triskelia interact they form a polyhedral lattice that surrounds the vesicle. The protein's name refers to this lattice structure, deriving from Latin clathri meaning lattice. Barbara Pearse named the protein clathrin at the suggestion of Graeme Mitchison, selecting it from three possible options. Coat-proteins, like clathrin, are used to build small vesicles in order to transport molecules within cells. The endocytosis and exocytosis of vesicles allows cells to communicate, to transfer nutrients, to import signaling receptors, to mediate an immune response after sampling the extracellular world, and to clean up the cell debris left by tissue inflammation. The endocytic pathway can be hijacked by viruses and other pathogens in order to gain entry to the cell during infection.
A kinetochore is a disc-shaped protein structure associated with duplicated chromatids in eukaryotic cells where the spindle fibers attach during cell division to pull sister chromatids apart. The kinetochore assembles on the centromere and links the chromosome to microtubule polymers from the mitotic spindle during mitosis and meiosis. The term kinetochore was first used in a footnote in a 1934 Cytology book by Lester W. Sharp and commonly accepted in 1936. Sharp's footnote reads: "The convenient term kinetochore has been suggested to the author by J. A. Moore", likely referring to John Alexander Moore who had joined Columbia University as a freshman in 1932.
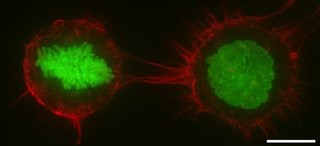
The cell cortex, also known as the actin cortex, cortical cytoskeleton or actomyosin cortex, is a specialized layer of cytoplasmic proteins on the inner face of the cell membrane. It functions as a modulator of membrane behavior and cell surface properties. In most eukaryotic cells lacking a cell wall, the cortex is an actin-rich network consisting of F-actin filaments, myosin motors, and actin-binding proteins. The actomyosin cortex is attached to the cell membrane via membrane-anchoring proteins called ERM proteins that plays a central role in cell shape control. The protein constituents of the cortex undergo rapid turnover, making the cortex both mechanically rigid and highly plastic, two properties essential to its function. In most cases, the cortex is in the range of 100 to 1000 nanometers thick.

Aurora kinase A also known as serine/threonine-protein kinase 6 is an enzyme that in humans is encoded by the AURKA gene.
An asymmetric cell division produces two daughter cells with different cellular fates. This is in contrast to symmetric cell divisions which give rise to daughter cells of equivalent fates. Notably, stem cells divide asymmetrically to give rise to two distinct daughter cells: one copy of the original stem cell as well as a second daughter programmed to differentiate into a non-stem cell fate.
Septins are a group of GTP-binding proteins expressed in all eukaryotic cells except plants. Different septins form protein complexes with each other. These complexes can further assemble into filaments, rings and gauzes. Assembled as such, septins function in cells by localizing other proteins, either by providing a scaffold to which proteins can attach, or by forming a barrier preventing the diffusion of molecules from one compartment of the cell to another, or in the cell cortex as a barrier to the diffusion of membrane-bound proteins.

In cell biology, a bleb is a bulge of the plasma membrane of a cell, characterized by a spherical, "blister-like", bulky morphology. It is characterized by the decoupling of the cytoskeleton from the plasma membrane, degrading the internal structure of the cell, allowing the flexibility required for the cell to separate into individual bulges or pockets of the intercellular matrix. Most commonly, blebs are seen in apoptosis but are also seen in other non-apoptotic functions. Blebbing, or zeiosis, is the formation of blebs.
In cell biology, microtubule nucleation is the event that initiates de novo formation of microtubules (MTs). These filaments of the cytoskeleton typically form through polymerization of α- and β-tubulin dimers, the basic building blocks of the microtubule, which initially interact to nucleate a seed from which the filament elongates.

Kinesin-like protein KIF2C is a protein that in humans is encoded by the KIF2C gene.

Cytoplasmic linker associated protein 1, also known as CLASP1, is a protein which in humans is encoded by the CLASP1 gene.

Anillin is a conserved protein implicated in cytoskeletal dynamics during cellularization and cytokinesis. The ANLN gene in humans and the scraps gene in Drosophila encode Anillin. In 1989, anillin was first isolated in embryos of Drosophila melanogaster. It was identified as an F-actin binding protein. Six years later, the anillin gene was cloned from cDNA originating from a Drosophila ovary. Staining with anti-anillin antibody showed the anillin localizes to the nucleus during interphase and to the contractile ring during cytokinesis. These observations agree with further research that found anillin in high concentrations near the cleavage furrow coinciding with RhoA, a key regulator of contractile ring formation.
Amitosis, also called karyostenosis, direct cell division or Binary fission, is a type of asexual cell division utilized by most prokaryotes. It differs from other forms of cell division as it does not involve the mitotic apparatus nor the condensation of chromatin into chromosomes.
Cell mechanics is a sub-field of biophysics that focuses on the mechanical properties and behavior of living cells and how it relates to cell function. It encompasses aspects of cell biophysics, biomechanics, soft matter physics and rheology, mechanobiology and cell biology.
Hertwig's rule, or the long axis rule states that a cell divides along its long axis. Introduced by the German zoologist Oscar Hertwig in 1884, the rule emphasizes the cell shape as a default mechanism of spindle apparatus orientation. Hertwig's rule predicts cell division orientation, which is important for tissue architecture, cell fate and morphogenesis.
Xenopus egg extract is a lysate that is prepared by crushing the eggs of the African clawed frog Xenopus laevis. It offers a powerful cell-free system for studying various cell biological processes, including cell cycle progression, nuclear transport, DNA replication and chromosome segregation. It is also called Xenopus egg cell-free system or Xenopus egg cell-free extract.
Barry James Thompson is an Australian and British developmental biologist and cancer biologist. Thompson is known for identifying genes, proteins and mechanisms involved in epithelial polarity, morphogenesis and cell signaling via the Wnt and Hippo signaling pathways, which have key roles in human cancer.
