
Sexual selection is a mode of natural selection in which members of one biological sex choose mates of the other sex to mate with, and compete with members of the same sex for access to members of the opposite sex. These two forms of selection mean that some individuals have greater reproductive success than others within a population, for example because they are more attractive or prefer more attractive partners to produce offspring. Successful males benefit from frequent mating and monopolizing access to one or more fertile females. Females can maximise the return on the energy they invest in reproduction by selecting and mating with the best males.

A lek is an aggregation of male animals gathered to engage in competitive displays and courtship rituals, known as lekking, to entice visiting females which are surveying prospective partners with which to mate. A lek can also indicate an available plot of space able to be utilized by displaying males to defend their own share of territory for the breeding season. A lekking species is characterised by male displays, strong female mate choice, and the conferring of indirect benefits to males and reduced costs to females. Although most prevalent among birds such as black grouse, lekking is also found in a wide range of vertebrates including some bony fish, amphibians, reptiles, and mammals, and arthropods including crustaceans and insects.

The handicap principle is a hypothesis proposed by the Israeli biologist Amotz Zahavi to explain how evolution may lead to "honest" or reliable signalling between animals which have an obvious motivation to bluff or deceive each other.
Fisherian runaway or runaway selection is a sexual selection mechanism proposed by the mathematical biologist Ronald Fisher in the early 20th century, to account for the evolution of ostentatious male ornamentation by persistent, directional female choice. An example is the colourful and elaborate peacock plumage compared to the relatively subdued peahen plumage; the costly ornaments, notably the bird's extremely long tail, appear to be incompatible with natural selection. Fisherian runaway can be postulated to include sexually dimorphic phenotypic traits such as behavior expressed by a particular sex.

Reproductive success is an individual's production of offspring per breeding event or lifetime. This is not limited by the number of offspring produced by one individual, but also the reproductive success of these offspring themselves.

The Platystomatidae are a distinctive family of flies (Diptera) in the superfamily Tephritoidea.

Mate choice is one of the primary mechanisms under which evolution can occur. It is characterized by a "selective response by animals to particular stimuli" which can be observed as behavior. In other words, before an animal engages with a potential mate, they first evaluate various aspects of that mate which are indicative of quality—such as the resources or phenotypes they have—and evaluate whether or not those particular trait(s) are somehow beneficial to them. The evaluation will then incur a response of some sort.

The sexy son hypothesis in evolutionary biology and sexual selection, proposed by Patrick J. Weatherhead and Raleigh J. Robertson of Queen's University in Kingston, Ontario in 1979, states that a female's ideal mate choice among potential mates is one whose genes will produce males with the best chance of reproductive success. This implies that other benefits the father can offer the mother or offspring are less relevant than they may appear, including his capacity as a parental caregiver, territory and any nuptial gifts. Fisher's principle means that the sex ratio is always near 1:1 between males and females, yet what matters most are her "sexy sons'" future breeding successes, more likely if they have a promiscuous father, in creating large numbers of offspring carrying copies of her genes. This sexual selection hypothesis has been researched in species such as the European pied flycatcher.
The lek paradox is the conundrum of how additive or beneficial genetic variation is maintained in lek mating species in the face of consistent sexual selection based on female preferences. While many studies have attempted to explain how the lek paradox fits into Darwinian theory, the paradox remains. Persistent female choice for particular male trait values should erode genetic diversity in male traits and thereby remove the benefits of choice, yet choice persists. This paradox can be somewhat alleviated by the occurrence of mutations introducing potential differences, as well as the possibility that traits of interest have more or less favorable recessive alleles.

A courtship display is a set of display behaviors in which an animal, usually a male, attempts to attract a mate; the mate exercises choice, so sexual selection acts on the display. These behaviors often include ritualized movement ("dances"), vocalizations, mechanical sound production, or displays of beauty, strength, or agonistic ability.

Many species have multiple sexual ornaments, whereby females select mating partners using several cues instead of only one cue. Whereas this phenomenon is self-evident and hence long recognized, adaptive explanations of why females use several instead of only one signal have been formulated relatively recently. Several hypotheses exist, but mutually exclusive tests are still lacking.
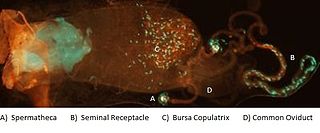
Female sperm storage is a biological process and often a type of sexual selection in which sperm cells transferred to a female during mating are temporarily retained within a specific part of the reproductive tract before the oocyte, or egg, is fertilized. This process takes place in some species of animals, but not in humans. The site of storage is variable among different animal taxa and ranges from structures that appear to function solely for sperm retention, such as insect spermatheca and bird sperm storage tubules, to more general regions of the reproductive tract enriched with receptors to which sperm associate before fertilization, such as the caudal portion of the cow oviduct containing sperm-associating annexins. Female sperm storage is an integral stage in the reproductive process for many animals with internal fertilization. It has several documented biological functions including:
A biological ornament is a characteristic of an animal that appears to serve a decorative function rather than a utilitarian function. Many are secondary sexual characteristics, and others appear on young birds during the period when they are dependent on being fed by their parents. Ornaments are used in displays to attract mates, which may lead to the evolutionary process known as sexual selection. An animal may shake, lengthen, or spread out its ornament in order to get the attention of the opposite sex, which will in turn choose the most attractive one with which to mate. Ornaments are most often observed in males, and choosing an extravagantly ornamented male benefits females as the genes that produce the ornament will be passed on to her offspring, increasing their own reproductive fitness. As Ronald Fisher noted, the male offspring will inherit the ornament while the female offspring will inherit the preference for said ornament, which can lead to a positive feedback loop known as a Fisherian runaway. These structures serve as cues to animal sexual behaviour, that is, they are sensory signals that affect mating responses. Therefore, ornamental traits are often selected by mate choice.
Phytalmia mouldsi is in the subfamily Phytalmiinae which includes all of the antlered fruit flies. They were discovered by M. S. Moulds in 1977 and are found only in an isolated rainforest of Australia. Antlered flies, collectively, are notable for their unique mating behavior and the unusual antler-like extensions on the heads of the males.
Interlocus sexual conflict is a type of sexual conflict that occurs through the interaction of a set of antagonistic alleles at two or more different loci, or the location of a gene on a chromosome, in males and females, resulting in the deviation of either or both sexes from the fitness optima for the traits. A co-evolutionary arms race is established between the sexes in which either sex evolves a set of antagonistic adaptations that is detrimental to the fitness of the other sex. The potential for reproductive success in one organism is strengthened while the fitness of the opposite sex is weakened. Interlocus sexual conflict can arise due to aspects of male–female interactions such as mating frequency, fertilization, relative parental effort, female remating behavior, and female reproductive rate.

Sepsis cynipsea is a European species of fly and member of the family Sepsidae. It is a coprophagous fly that feeds on dung. These flies are most commonly found around freshly laid cattle dung where they eat and reproduce. Due to human agricultural practices involving cows, these flies are now common in other areas of the world.

Prochyliza xanthostoma, the waltzing fly, is a species of carrion-feeding cheese skipper, insects in the family Piophilidae and the order Diptera. P. xanthostoma is a member of the genus Prochyliza, which contains eleven species. The adult flies are found through North America and are brown-bodied, with orange and black coloring. Mating occurs on animal carcasses and male perform mating rituals; females engage in ejaculate feeding. The waltzing fly is known for its exaggerated sexual dimorphism and has thus become a prominent model for sexual dimorphism and larval behavior. These organisms are known as cheese skippers because when startled, the larvae can leap several inches into the air. P. xanthostoma is an important model organism for sexual selection, larval behavior, and adult reproductive success and survivability.
Derocephalus angusticollis is a fly in the family Neriidae. They are typically found on the east coast of Australia near rotting vegetation. Aggregating on the rotting bark of trees such as Acacia longifolia and other trees in New South Wales and southern Queensland. D. angusticollis flies found in the wild have accelerated speeds of development and age of mortality when compared to those in captivity. Derocephalus One characteristic of the neriid fly is that it demonstrates sexual dimorphism. Males have a larger build as well as exaggerated physical characteristics such as wider heads and longer limbs. Certain phenotypic characteristics are dependent on the diet of the parents.

Teleopsis dalmanni, synonym Cyrtodiopsis dalmanni, also known as the Malaysian stalk-eyed fly, is a species of fly in the family Diopsidae. T. dalmanni flies possess lateral elongations on their head capsules called eyestalks. These eyestalks play an important role in mate selection and as a result physical characteristic of the fly has been the subject of several studies on sexual selection, natural selection, and mating behavior.

Richardia telescopica is a species of flies from the genus Richardia. The species was originally described by Gerstaecker in 1860 and it occurs in Nicaragua, Costa Rica, and Panama.



