Sex is the trait that determines whether a sexually reproducing organism produces male or female gametes. During sexual reproduction, a male and a female gamete fuse to form a zygote, which develops into an offspring that inherits traits from each parent. By convention, organisms that produce smaller, more mobile gametes are called male, while organisms that produce produce larger, non-mobile gametes are called female. An organism that produces both types of gamete is hermaphrodite.
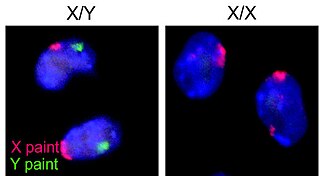
The XY sex-determination system is a sex-determination system used to classify many mammals, including humans, some insects (Drosophila), some snakes, some fish (guppies), and some plants. In this system, the sex of an individual is determined by a pair of sex chromosomes. In most cases, females have two of the same kind of sex chromosome (XX), and are called the homogametic sex. Males have two different kinds of sex chromosomes (XY), and are called the heterogametic sex.

A sex-determination system is a biological system that determines the development of sexual characteristics in an organism. Most organisms that create their offspring using sexual reproduction have two common sexes and a few less common intersex variations.

In animals, viviparity is development of the embryo inside the body of the mother, with the maternal circulation providing for the metabolic needs of the embryo's development, until the mother gives birth to a fully or partially developed juvenile that is at least metabolically independent. This is opposed to oviparity, where the embryos develop independently outside the mother in eggs until they are developed enough to break out as hatchlings; and ovoviviparity, where the embryos are developed in eggs that remain carried inside the mother's body until the hatchlings emerge from the mother as juveniles, similar to a live birth.
A sex ratio is the ratio of males to females in a population. As explained by Fisher's principle, for evolutionary reasons this is typically about 1:1 in species which reproduce sexually. However, many species deviate from an even sex ratio, either periodically or permanently. Examples include parthenogenic species, periodically mating organisms such as aphids, some eusocial wasps, bees, ants, and termites.

Sexual differentiation is the process of development of the sex differences between males and females from an undifferentiated zygote. Sex determination is often distinct from sex differentiation; sex determination is the designation for the development stage towards either male or female, while sex differentiation is the pathway towards the development of the phenotype.

The spotted turtle, the only species of the genus Clemmys, is a small, semi-aquatic turtle that reaches a carapace length of 8–12 cm (3.1–4.7 in) upon adulthood. Their broad, smooth, low dark-colored upper shell, or carapace, ranges in its exact colour from black to a bluish black with a number of tiny yellow round spots. The spotting patterning extends from the head, to the neck and out onto the limbs. Sexually mature males have a concave plastron and a long, thick tail. By contrast, sexually mature females possess a flat plastron and have a tail that is noticeably shorter and thinner than that of mature males. Mature males also have a dark iris and face; females typically have a yellow or orange iris and a similarly coloured face that is distinctly lighter than the males'. Juveniles appear female-like in this regard, and at maturity males begin to develop darker features.

Aromatase, also called estrogen synthetase or estrogen synthase, is an enzyme responsible for a key step in the biosynthesis of estrogens. It is CYP19A1, a member of the cytochrome P450 superfamily, which are monooxygenases that catalyze many reactions involved in steroidogenesis. In particular, aromatase is responsible for the aromatization of androgens into estrogens. The enzyme aromatase can be found in many tissues including gonads, brain, adipose tissue, placenta, blood vessels, skin, and bone, as well as in tissue of endometriosis, uterine fibroids, breast cancer, and endometrial cancer. It is an important factor in sexual development.

A polyphenic trait is a trait for which multiple, discrete phenotypes can arise from a single genotype as a result of differing environmental conditions. It is therefore a special case of phenotypic plasticity.

The XO sex-determination system is a system that some species of insects, arachnids, and mammals use to determine the sex of offspring. In this system, there is only one sex chromosome, referred to as X. Males only have one X chromosome (XO), while females have two (XX). The letter O signifies the lack of a Y chromosome. Maternal gametes always contain an X chromosome, so the sex of the animals' offspring depends on whether a sex chromosome is present in the male gamete. Its sperm normally contains either one X chromosome or no sex chromosomes at all.
The ZW sex-determination system is a chromosomal system that determines the sex of offspring in birds, some fish and crustaceans such as the giant river prawn, some insects, the schistosome family of flatworms, and some reptiles, e.g. majority of snakes, lacertid lizards and monitors, including Komodo dragons. It is also present in some plants, where it has probably evolved independently on several occasions. The letters Z and W are used to distinguish this system from the XY sex-determination system. In the ZW system, females have a pair of dissimilar ZW chromosomes, and males have two similar ZZ chromosomes.
Sex allocation is the allocation of resources to male versus female reproduction in sexual species. Sex allocation theory tries to explain why many species produce equal number of males and females.
Sex chromosomes are chromosomes that carry the genes that determine the sex of an individual. The human sex chromosomes are a typical pair of mammal allosomes. They differ from autosomes in form, size, and behavior. Whereas autosomes occur in homologous pairs whose members have the same form in a diploid cell, members of an allosome pair may differ from one another.

Environmental sex determination is the establishment of sex by a non-genetic cue, such as nutrient availability, experienced within a discrete period after fertilization. Environmental factors which often influence sex determination during development or sexual maturation include light intensity and photoperiod, temperature, nutrient availability, and pheromones emitted by surrounding plants or animals. This is in contrast to genotypic sex determination, which establishes sex at fertilization by genetic factors such as sex chromosomes. Under true environmental sex determination, once sex is determined, it is fixed and cannot be switched again. Environmental sex determination is different from some forms of sequential hermaphroditism in which the sex is determined flexibly after fertilization throughout the organism’s life.
Fisher's principle is an evolutionary model that explains why the sex ratio of most species that produce offspring through sexual reproduction is approximately 1:1 between males and females. A. W. F. Edwards has remarked that it is "probably the most celebrated argument in evolutionary biology".

The eastern three-lined skink, also known commonly as the bold-striped cool-skink, is a species of skink, a lizard in the family Scincidae. The species is endemic to Australia. A. duperreyi has been extensively studied in the context of understanding the evolution of learning, viviparity in lizards, and temperature- and genetic-sex determination. A. duperreyi is classified as a species of "Least Concern" by the IUCN.
Sex reversal is a biological process whereby the pathway directed towards the already determined-sex fate is flipped towards the opposite sex, creating a discordance between the primary sex fate and the sex phenotype expressed. The process of sex reversal occurs during embryonic development or before gonad differentiation. In GSD species, sex reversal means that the sexual phenotype is discordant with the genetic/chromosomal sex. In TSD species, sex reversal means that the temperature/conditions that usually trigger the differentiation towards one sexual phenotype are producing the opposite sexual phenotype.

David Pafford Crews is the Ashbel Smith Professor of Zoology and Psychology at the University of Texas at Austin. He has been a pioneer in several areas of reproductive biology, including evolution of sexual behavior and differentiation, neural and phenotypic plasticity, and the role of endocrine disruptors on brain and behavior.
Ecological evolutionary developmental biology (eco-evo-devo) is a field of biology combining ecology, developmental biology and evolutionary biology to examine their relationship. The concept is closely tied to multiple biological mechanisms. The effects of eco-evo-devo can be a result of developmental plasticity, the result of symbiotic relationships or epigenetically inherited. The overlap between developmental plasticity and symbioses rooted in evolutionary concepts defines ecological evolutionary developmental biology. Host- microorganisms interactions during development characterize symbiotic relationships, whilst the spectrum of phenotypes rooted in canalization with response to environmental cues highlights plasticity. Developmental plasticity that is controlled by environmental temperature may put certain species at risk as a result of climate change.

The evolution of sex-determining mechanisms, characterized by the evolutionary transition to genetic sex determination or temperature-dependent sex determination from the opposite mechanism, has frequently and readily occurred among multiple taxa across a transitionary continuum.