Related Research Articles

The immune system is a network of biological systems that protects an organism from diseases. It detects and responds to a wide variety of pathogens, from viruses to parasitic worms, as well as cancer cells and objects such as wood splinters, distinguishing them from the organism's own healthy tissue. Many species have two major subsystems of the immune system. The innate immune system provides a preconfigured response to broad groups of situations and stimuli. The adaptive immune system provides a tailored response to each stimulus by learning to recognize molecules it has previously encountered. Both use molecules and cells to perform their functions.

Macrophages are a type of white blood cell of the innate immune system that engulf and digest pathogens, such as cancer cells, microbes, cellular debris, and foreign substances, which do not have proteins that are specific to healthy body cells on their surface. This process is called phagocytosis, which acts to defend the host against infection and injury.

Phagocytosis is the process by which a cell uses its plasma membrane to engulf a large particle, giving rise to an internal compartment called the phagosome. It is one type of endocytosis. A cell that performs phagocytosis is called a phagocyte.

Neutrophils are a type of white blood cell. More specifically, they form the most abundant type of granulocytes and make up 40% to 70% of all white blood cells in humans. They form an essential part of the innate immune system, with their functions varying in different animals.
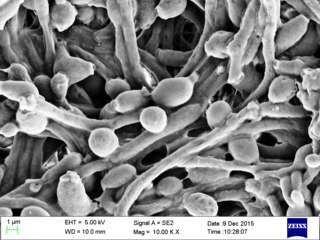
Candida albicans is an opportunistic pathogenic yeast that is a common member of the human gut flora. It can also survive outside the human body. It is detected in the gastrointestinal tract and mouth in 40–60% of healthy adults. It is usually a commensal organism, but it can become pathogenic in immunocompromised individuals under a variety of conditions. It is one of the few species of the genus Candida that cause the human infection candidiasis, which results from an overgrowth of the fungus. Candidiasis is, for example, often observed in HIV-infected patients. C. albicans is the most common fungal species isolated from biofilms either formed on (permanent) implanted medical devices or on human tissue. C. albicans, C. tropicalis, C. parapsilosis, and C. glabrata are together responsible for 50–90% of all cases of candidiasis in humans. A mortality rate of 40% has been reported for patients with systemic candidiasis due to C. albicans. By one estimate, invasive candidiasis contracted in a hospital causes 2,800 to 11,200 deaths yearly in the US. Nevertheless, these numbers may not truly reflect the true extent of damage this organism causes, given new studies indicating that C. albicans can cross the blood–brain barrier in mice.
Phagocytes are cells that protect the body by ingesting harmful foreign particles, bacteria, and dead or dying cells. Their name comes from the Greek phagein, "to eat" or "devour", and "-cyte", the suffix in biology denoting "cell", from the Greek kutos, "hollow vessel". They are essential for fighting infections and for subsequent immunity. Phagocytes are important throughout the animal kingdom and are highly developed within vertebrates. One litre of human blood contains about six billion phagocytes. They were discovered in 1882 by Ilya Ilyich Mechnikov while he was studying starfish larvae. Mechnikov was awarded the 1908 Nobel Prize in Physiology or Medicine for his discovery. Phagocytes occur in many species; some amoebae behave like macrophage phagocytes, which suggests that phagocytes appeared early in the evolution of life.

Cryptococcus neoformans is an encapsulated yeast belonging to the class Tremellomycetes and an obligate aerobe that can live in both plants and animals. Its teleomorph is a filamentous fungus, formerly referred to Filobasidiella neoformans. In its yeast state, it is often found in bird excrement. Cryptococcus neoformans can cause disease in apparently immunocompetent, as well as immunocompromised, hosts.

In cell biology, a phagosome is a vesicle formed around a particle engulfed by a phagocyte via phagocytosis. Professional phagocytes include macrophages, neutrophils, and dendritic cells (DCs).
Intracellular parasites are microparasites that are capable of growing and reproducing inside the cells of a host.

The innate, or nonspecific, immune system is one of the two main immunity strategies in vertebrates. The innate immune system is an alternate defense strategy and is the dominant immune system response found in plants, fungi, insects, and primitive multicellular organisms.

In biology, a phagolysosome, or endolysosome, is a cytoplasmic body formed by the fusion of a phagosome with a lysosome in a process that occurs during phagocytosis. Formation of phagolysosomes is essential for the intracellular destruction of microorganisms and pathogens. It takes place when the phagosome's and lysosome's membranes 'collide', at which point the lysosomal contents—including hydrolytic enzymes—are discharged into the phagosome in an explosive manner and digest the particles that the phagosome had ingested. Some products of the digestion are useful materials and are moved into the cytoplasm; others are exported by exocytosis.

An alveolar macrophage, pulmonary macrophage, is a type of macrophage, a professional phagocyte, found in the airways and at the level of the alveoli in the lungs, but separated from their walls.
The mannose receptor is a C-type lectin primarily present on the surface of macrophages, immature dendritic cells and liver sinusoidal endothelial cells, but is also expressed on the surface of skin cells such as human dermal fibroblasts and keratinocytes. It is the first member of a family of endocytic receptors that includes Endo180 (CD280), M-type PLA2R, and DEC-205 (CD205).
Pathogenic fungi are fungi that cause disease in humans or other organisms. Although fungi are eukaryotic, many pathogenic fungi are microorganisms. Approximately 300 fungi are known to be pathogenic to humans; their study is called "medical mycology". Fungal infections kill more people than either tuberculosis or malaria—about 2 million people per year.

A blastoconidium is an asexual holoblastic conidia formed through the blowing out or budding process of a yeast cell, which is a type of asexual reproduction that results in a bud arising from a parent cell. The production of a blastoconidium can occur along a true hyphae, pseudohyphae, or a singular yeast cell. The word "conidia" comes from the Greek word konis and eidos, konis meaning dust and eidos meaning like. The term "bud" comes from the Greek word blastos, which means bud. Yeasts such as Candida albicans and Cryptococcus neoformans produce these budded cells known as blastoconidia.

Immunity Related Guanosine Triphosphatases or IRGs are proteins activated as part of an early immune response. IRGs have been described in various mammals but are most well characterized in mice. IRG activation in most cases is induced by an immune response and leads to clearance of certain pathogens.
Membrane vesicle trafficking in eukaryotic animal cells involves movement of biochemical signal molecules from synthesis-and-packaging locations in the Golgi body to specific release locations on the inside of the plasma membrane of the secretory cell. It takes place in the form of Golgi membrane-bound micro-sized vesicles, termed membrane vesicles (MVs).

Arturo Casadevall is a Bloomberg Distinguished Professor of Molecular Microbiology & Immunology and Infectious Diseases at the Johns Hopkins Bloomberg School of Public Health and Johns Hopkins School of Medicine, and the Alfred and Jill Sommer Professor and Chair of the W. Harry Feinstone Department of Molecular Microbiology and Immunology at the Johns Hopkins Bloomberg School of Public Health. He is an internationally recognized expert in infectious disease research, with a focus on fungal and bacterial pathogenesis and basic immunology of antibody structure-function. He was elected a member of the National Academy of Sciences in 2022.
Candidalysin is a cytolytic 31-amino acid α-helical amphipathic peptide toxin secreted by the opportunistic pathogen Candida albicans. This toxin is a fungal example of a classical virulence factor. Hyphal morphogenesis in C. albicans is associated with damage to host epithelial cells; during this process Candidalysin is released and intercalates in host membranes. Candidalysin promotes damage of oral epithelial cells and induces lactate dehydrogenase release and calcium ion influx. It is unique in the fact that it is the first peptide toxin to be identified in any human fungal pathogen.

Joseph Heitman is an American physician-scientist focused on research in genetics, microbiology, and infectious diseases. He is the James B. Duke Professor and Chair of the Department of Molecular Genetics and Microbiology at Duke University School of Medicine.
References
- ↑ Ma H, Croudace JE, Lammas DA, May RC (November 2006). "Expulsion of live pathogenic yeast by macrophages". Current Biology. 16 (21): 2156–60. doi: 10.1016/j.cub.2006.09.032 . PMID 17084701. S2CID 11639313.
- ↑ Alvarez M, Casadevall A (November 2006). "Phagosome extrusion and host-cell survival after Cryptococcus neoformans phagocytosis by macrophages". Current Biology. 16 (21): 2161–5. doi: 10.1016/j.cub.2006.09.061 . PMID 17084702. S2CID 1612240.
- ↑ Bain JM, Lewis LE, Okai B, Quinn J, Gow NA, Erwig LP (September 2012). "Non-lytic expulsion/exocytosis of Candida albicans from macrophages". Fungal Genetics and Biology. 49 (9): 677–8. doi:10.1016/j.fgb.2012.01.008. PMC 3430864 . PMID 22326419.
- ↑ García-Rodas R, González-Camacho F, Rodríguez-Tudela JL, Cuenca-Estrella M, Zaragoza O (June 2011). "The interaction between Candida krusei and murine macrophages results in multiple outcomes, including intracellular survival and escape from killing". Infection and Immunity. 79 (6): 2136–44. doi:10.1128/iai.00044-11. PMC 3125833 . PMID 21422181.
- ↑ Johnston SA, May RC (March 2013). "Cryptococcus interactions with macrophages: evasion and manipulation of the phagosome by a fungal pathogen". Cellular Microbiology. 15 (3): 403–11. doi:10.1111/cmi.12067. PMID 23127124. S2CID 39991842.
- ↑ Hagedorn M, Rohde KH, Russell DG, Soldati T (March 2009). "Infection by tubercular mycobacteria is spread by nonlytic ejection from their amoeba hosts". Science. 323 (5922): 1729–33. Bibcode:2009Sci...323.1729H. doi:10.1126/science.1169381. PMC 2770343 . PMID 19325115.
- ↑ Johnston SA, Voelz K, May RC (February 2016). "Cryptococcus neoformans Thermotolerance to Avian Body Temperature Is Sufficient For Extracellular Growth But Not Intracellular Survival In Macrophages". Scientific Reports. 6: 20977. Bibcode:2016NatSR...620977J. doi:10.1038/srep20977. PMC 4756366 . PMID 26883088.
- ↑ Bojarczuk A, Miller KA, Hotham R, Lewis A, Ogryzko NV, Kamuyango AA, et al. (February 2016). "Cryptococcus neoformans Intracellular Proliferation and Capsule Size Determines Early Macrophage Control of Infection". Scientific Reports. 6: 21489. Bibcode:2016NatSR...621489B. doi:10.1038/srep21489. PMC 4757829 . PMID 26887656.
- ↑ Nicola AM, Robertson EJ, Albuquerque P, Derengowski LD, Casadevall A (2011). "Nonlytic exocytosis of Cryptococcus neoformans from macrophages occurs in vivo and is influenced by phagosomal pH". mBio. 2 (4): e00167-11. doi:10.1128/mBio.00167-11. PMC 3150755 . PMID 21828219.
- ↑ Watkins RA, Andrews A, Wynn C, Barisch C, King JS, Johnston SA (April 9, 2018). "Cryptococcus neoformans Escape From Dictyostelium Amoeba by Both WASH-Mediated Constitutive Exocytosis and Vomocytosis". Frontiers in Cellular and Infection Microbiology. 8 (108): 108. doi: 10.3389/fcimb.2018.00108 . PMC 5900056 . PMID 29686972.
- 1 2 3 4 5 6 7 8 9 Seoane PI, May RC (February 2020). "Vomocytosis: What we know so far". Cellular Microbiology. 22 (2): e13145. doi: 10.1111/cmi.13145 . PMID 31730731. S2CID 208061582.
- 1 2 3 4 5 Cruz-Acuña M, Pacifici N, Lewis JS (December 2019). Garsin DA (ed.). "Vomocytosis: Too Much Booze, Base, or Calcium?". mBio. 10 (6): e02526–19, /mbio/10/6/mBio.02526–19.atom. doi:10.1128/mBio.02526-19. PMC 6935858 . PMID 31874916.
- ↑ Lundie RJ, Helbig KJ, Pearson JS, Fairfax KA (January 2019). "Fluorescent antibiotics, vomocytosis, vaccine candidates and the inflammasome". Clinical & Translational Immunology. 8 (11): e01083. doi:10.1002/cti2.1083. PMC 6823609 . PMID 31700626.
- ↑ Voelz K, Lammas DA, May RC (August 2009). "Cytokine signaling regulates the outcome of intracellular macrophage parasitism by Cryptococcus neoformans". Infection and Immunity. 77 (8): 3450–7. doi:10.1128/iai.00297-09. PMC 2715691 . PMID 19487474.
- ↑ Johnston SA, May RC (August 2010). "The human fungal pathogen Cryptococcus neoformans escapes macrophages by a phagosome emptying mechanism that is inhibited by Arp2/3 complex-mediated actin polymerisation". PLOS Pathogens. 6 (8): e1001041. doi: 10.1371/journal.ppat.1001041 . PMC 2920849 . PMID 20714349.
- 1 2 Gilbert AS, Seoane PI, Sephton-Clark P, Bojarczuk A, Hotham R, Giurisato E, et al. (August 2017). "Vomocytosis of live pathogens from macrophages is regulated by the atypical MAP kinase ERK5". Science Advances. 3 (8): e1700898. Bibcode:2017SciA....3E0898G. doi:10.1126/sciadv.1700898. PMC 5559206 . PMID 28835924.
- ↑ Alvarez M, Casadevall A (August 2007). "Cell-to-cell spread and massive vacuole formation after Cryptococcus neoformans infection of murine macrophages". BMC Immunology. 8: 16. doi: 10.1186/1471-2172-8-16 . PMC 1988836 . PMID 17705844.
- ↑ Sharma NK, Sarode SC, Sarode GS, Patil S (May 2019). "Vomocytosis by macrophages: a crucial event in the local niche of tumors". Future Oncology. 15 (14): 1545–1550. doi: 10.2217/fon-2019-0078 . PMID 31038349.