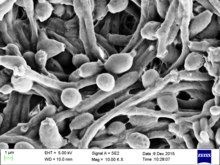
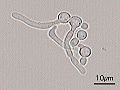
Candidiasis is a fungal infection due to any species of the genus Candida. When it affects the mouth, in some countries it is commonly called thrush. Signs and symptoms include white patches on the tongue or other areas of the mouth and throat. Other symptoms may include soreness and problems swallowing. When it affects the vagina, it may be referred to as a yeast infection or thrush. Signs and symptoms include genital itching, burning, and sometimes a white "cottage cheese-like" discharge from the vagina. Yeast infections of the penis are less common and typically present with an itchy rash. Very rarely, yeast infections may become invasive, spreading to other parts of the body. This may result in fevers, among other symptoms.
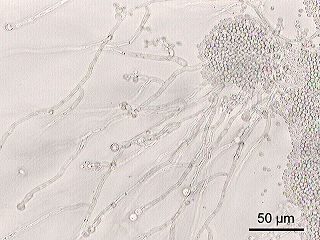
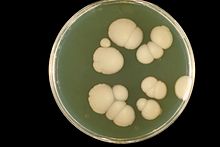
Candida is a genus of yeasts. It is the most common cause of fungal infections worldwide and the largest genus of medically important yeast.
Oral candidiasis (Acute pseudomembranous candidiasis), also known as oral thrush among other names, is candidiasis that occurs in the mouth. That is, oral candidiasis is a mycosis (yeast/fungal infection) of Candida species on the mucous membranes of the mouth.
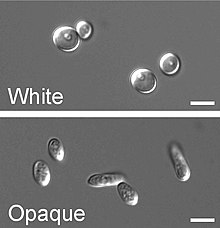
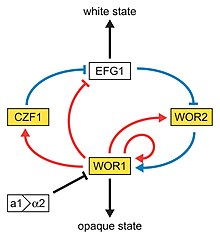
Phenotypic switching is switching between multiple cellular morphologies. David R. Soll described two such systems: the first high frequency switching system between several morphological stages and a second high frequency switching system between opaque and white cells. The latter is an epigenetic switching system

Terconazole is an antifungal drug used to treat vaginal yeast infection. It comes as a lotion or a suppository and disrupts the biosynthesis of fats in a yeast cell. It has a relatively broad spectrum compared to azole compounds but not triazole compounds. Testing shows that it is a suitable compound for prophylaxis for those that suffer from chronic vulvovaginal candidiasis.
Candida parapsilosis is a fungal species of yeast that has become a significant cause of sepsis and of wound and tissue infections in immunocompromised people. Unlike Candida albicans and Candida tropicalis, C. parapsilosis is not an obligate human pathogen, having been isolated from nonhuman sources such as domestic animals, insects and soil. C. parapsilosis is also a normal human commensal and it is one of the fungi most frequently isolated from human hands. There are several risk factors that can contribute to C. parapsilosis colonization. Immunocompromised individuals and surgical patients, particularly those undergoing surgery of the gastrointestinal tract, are at high risk for infection with C. parapsilosis. There is currently no consensus on the treatment of invasive candidiasis caused by C. parapsilosis, although the therapeutic approach typically includes the removal of foreign bodies such as implanted prostheses and the administration of systemic antifungal therapy. Amphotericin B and Fluconazole are often used in the treatment of C. parapsilosis infection.

Fungi are a diverse group of organisms that employ a huge variety of reproductive strategies, ranging from fully asexual to almost exclusively sexual species. Most species can reproduce both sexually and asexually, alternating between haploid and diploid forms. This contrasts with most multicellular eukaryotes such as mammals, where the adults are usually diploid and produce haploid gametes which combine to form the next generation. In fungi, both haploid and diploid forms can reproduce – haploid individuals can undergo asexual reproduction while diploid forms can produce gametes that combine to give rise to the next generation.
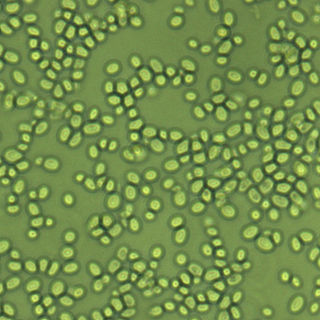
Nakaseomyces glabratus is a species of haploid yeast of the genus Nakaseomyces, previously known as Candida glabrata. Despite the fact that no sexual life cycle has been documented for this species, N. glabratus strains of both mating types are commonly found. N. glabrata is generally a commensal of human mucosal tissues, but in today's era of wider human immunodeficiency from various causes, N. glabratus is often the second or third most common cause of candidiasis as an opportunistic pathogen. Infections caused by N. glabratus can affect the urogenital tract or even cause systemic infections by entrance of the fungal cells in the bloodstream (Candidemia), especially prevalent in immunocompromised patients.

Dimorphic fungi are fungi that can exist in the form of both mold and yeast. This is usually brought about by change in temperature and the fungi are also described as thermally dimorphic fungi. An example is Talaromyces marneffei, a human pathogen that grows as a mold at room temperature, and as a yeast at human body temperature.
Pathogenic fungi are fungi that cause disease in humans or other organisms. Although fungi are eukaryotic, many pathogenic fungi are microorganisms. Approximately 300 fungi are known to be pathogenic to humans; their study is called "medical mycology". Fungal infections are estimated to kill more people than either tuberculosis or malaria—about two million people per year.

A blastoconidium is an asexual holoblastic conidia formed through the blowing out or budding process of a yeast cell, which is a type of asexual reproduction that results in a bud arising from a parent cell. The production of a blastoconidium can occur along a true hyphae, pseudohyphae, or a singular yeast cell. The word "conidia" comes from the Greek word konis and eidos, konis meaning dust and eidos meaning like. The term "bud" comes from the Greek word blastos, which means bud. Yeasts such as Candida albicans and Cryptococcus neoformans produce these budded cells known as blastoconidia.
Persister cells are subpopulations of cells that resist treatment, and become antimicrobial tolerant by changing to a state of dormancy or quiescence. Persister cells in their dormancy do not divide. The tolerance shown in persister cells differs from antimicrobial resistance in that the tolerance is not inherited and is reversible. When treatment has stopped the state of dormancy can be reversed and the cells can reactivate and multiply. Most persister cells are bacterial, and there are also fungal persister cells, yeast persister cells, and cancer persister cells that show tolerance for cancer drugs.
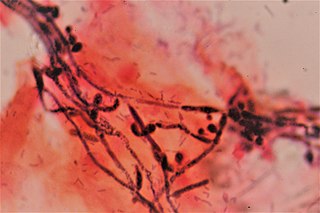
Vaginal yeast infection, also known as candidal vulvovaginitis and vaginal thrush, is excessive growth of yeast in the vagina that results in irritation. The most common symptom is vaginal itching, which may be severe. Other symptoms include burning with urination, a thick, white vaginal discharge that typically does not smell bad, pain during sex, and redness around the vagina. Symptoms often worsen just before a woman's period.

David R. Soll is a professor of Biology at the University of Iowa. He is best known for the motion analysis of living cells, the discovery of Candida albicans phenotypic switching and monoclonal antibody technology.
Mycobiota are a group of all the fungi present in a particular geographic region or habitat type. An analogous term for Mycobiota is funga.

Hwp1 is a protein (glycoprotein) located on the surface of an opportunistic diploid fungus called Candida albicans.
Fungal adhesins are proteins located on the surface of fungal cells, specifically found on the outside of the cell wall. They allow fungi to colonize various substrates and to bind to host tissues. Adhesion to tissue is an obligatory first step in pathogenesis by many yeasts. Adhesins also have other functions, such as mating and biofilm formation.

Candida auris is a species of fungus that grows as yeast. It is one of the few species of the genus Candida which cause candidiasis in humans. Often, candidiasis is acquired in hospitals by patients with weakened immune systems. C. auris can cause invasive candidiasis (fungemia) in which the bloodstream, the central nervous system, and internal organs are infected. It has attracted widespread attention because of its multiple drug resistance. Treatment is also complicated because it is easily misidentified as other Candida species.
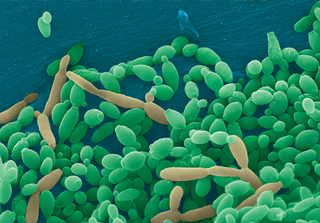
Candida tropicalis is a species of yeast in the genus Candida. It is a common pathogen in neutropenic hosts, in whom it may spread through the bloodstream to peripheral organs. For invasive disease, treatments include amphotericin B, echinocandins, or extended-spectrum triazole antifungals.
Carol Kumamoto is an American microbiologist who is Professor of Molecular Biology & Microbiology at Tufts University. She investigates the filamentous growth of Candida albicans, a fungal pathogen that causes several diseases. She is also interested in how C. albicans interacts with its host during colonisation and invasive diseases. She is a Fellow of the American Association for the Advancement of Science and the American Academy of Microbiology.