Related Research Articles

Meiosis is a special type of cell division of germ cells in sexually-reproducing organisms that produces the gametes, the sperm or egg cells. It involves two rounds of division that ultimately result in four cells, each with only one copy of each chromosome (haploid). Additionally, prior to the division, genetic material from the paternal and maternal copies of each chromosome is crossed over, creating new combinations of code on each chromosome. Later on, during fertilisation, the haploid cells produced by meiosis from a male and a female will fuse to create a zygote, a cell with two copies of each chromosome again.

Ploidy is the number of complete sets of chromosomes in a cell, and hence the number of possible alleles for autosomal and pseudoautosomal genes. Sets of chromosomes refer to the number of maternal and paternal chromosome copies, respectively, in each homologous chromosome pair, which chromosomes naturally exist as. Somatic cells, tissues, and individual organisms can be described according to the number of sets of chromosomes present : monoploid, diploid, triploid, tetraploid, pentaploid, hexaploid, heptaploid or septaploid, etc. The generic term polyploid is often used to describe cells with three or more sets of chromosomes.

A zygote is a eukaryotic cell formed by a fertilization event between two gametes. The zygote's genome is a combination of the DNA in each gamete, and contains all of the genetic information of a new individual organism. The sexual fusion of haploid cells is called karyogamy, the result of which is the formation of a diploid cell called the zygote or zygospore.

Basidiomycota is one of two large divisions that, together with the Ascomycota, constitute the subkingdom Dikarya within the kingdom Fungi. Members are known as basidiomycetes. More specifically, Basidiomycota includes these groups: agarics, puffballs, stinkhorns, bracket fungi, other polypores, jelly fungi, boletes, chanterelles, earth stars, smuts, bunts, rusts, mirror yeasts, and Cryptococcus, the human pathogenic yeast.

Ascomycota is a phylum of the kingdom Fungi that, together with the Basidiomycota, forms the subkingdom Dikarya. Its members are commonly known as the sac fungi or ascomycetes. It is the largest phylum of Fungi, with over 64,000 species. The defining feature of this fungal group is the "ascus", a microscopic sexual structure in which nonmotile spores, called ascospores, are formed. However, some species of Ascomycota are asexual and thus do not form asci or ascospores. Familiar examples of sac fungi include morels, truffles, brewers' and bakers' yeast, dead man's fingers, and cup fungi. The fungal symbionts in the majority of lichens such as Cladonia belong to the Ascomycota.
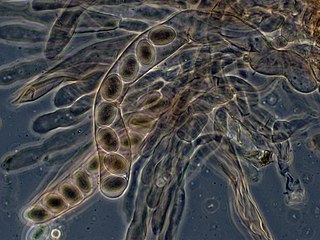
An ascus is the sexual spore-bearing cell produced in ascomycete fungi. Each ascus usually contains eight ascospores, produced by meiosis followed, in most species, by a mitotic cell division. However, asci in some genera or species can occur in numbers of one, two, four, or multiples of four. In a few cases, the ascospores can bud off conidia that may fill the asci with hundreds of conidia, or the ascospores may fragment, e.g. some Cordyceps, also filling the asci with smaller cells. Ascospores are nonmotile, usually single celled, but not infrequently may be coenocytic, and in some cases coenocytic in multiple planes. Mitotic divisions within the developing spores populate each resulting cell in septate ascospores with nuclei. The term ocular chamber, or oculus, refers to the epiplasm that is surrounded by the "bourrelet".
Gametogenesis is a biological process by which diploid or haploid precursor cells undergo cell division and differentiation to form mature haploid gametes. Depending on the biological life cycle of the organism, gametogenesis occurs by meiotic division of diploid gametocytes into various gametes, or by mitosis. For example, plants produce gametes through mitosis in gametophytes. The gametophytes grow from haploid spores after sporic meiosis. The existence of a multicellular, haploid phase in the life cycle between meiosis and gametogenesis is also referred to as alternation of generations.

In biology, a biological life cycle is a series of stages of the life of an organism, that begins as a zygote, often in an egg, and concludes as an adult that reproduces, producing an offspring in the form of a new zygote which then itself goes through the same series of stages, the process repeating in a cyclic fashion.

Karyogamy is the final step in the process of fusing together two haploid eukaryotic cells, and refers specifically to the fusion of the two nuclei. Before karyogamy, each haploid cell has one complete copy of the organism's genome. In order for karyogamy to occur, the cell membrane and cytoplasm of each cell must fuse with the other in a process known as plasmogamy. Once within the joined cell membrane, the nuclei are referred to as pronuclei. Once the cell membranes, cytoplasm, and pronuclei fuse, the resulting single cell is diploid, containing two copies of the genome. This diploid cell, called a zygote or zygospore can then enter meiosis, or continue to divide by mitosis. Mammalian fertilization uses a comparable process to combine haploid sperm and egg cells (gametes) to create a diploid fertilized egg.

Neurospora crassa is a type of red bread mold of the phylum Ascomycota. The genus name, meaning 'nerve spore' in Greek, refers to the characteristic striations on the spores. The first published account of this fungus was from an infestation of French bakeries in 1843.
Heterothallic species have sexes that reside in different individuals. The term is applied particularly to distinguish heterothallic fungi, which require two compatible partners to produce sexual spores, from homothallic ones, which are capable of sexual reproduction from a single organism.

A heterokaryon is a multinucleate cell that contains genetically different nuclei. Heterokaryotic and heterokaryosis are derived terms. This is a special type of syncytium. This can occur naturally, such as in the mycelium of fungi during sexual reproduction, or artificially as formed by the experimental fusion of two genetically different cells, as e.g., in hybridoma technology.
An oogonium is a small diploid cell which, upon maturation, forms a primordial follicle in a female fetus or the female gametangium of certain thallophytes.

Aspergillus nidulans is one of many species of filamentous fungi in the phylum Ascomycota. It has been an important research organism for studying eukaryotic cell biology for over 50 years, being used to study a wide range of subjects including recombination, DNA repair, mutation, cell cycle control, tubulin, chromatin, nucleokinesis, pathogenesis, metabolism, and experimental evolution. It is one of the few species in its genus able to form sexual spores through meiosis, allowing crossing of strains in the laboratory. A. nidulans is a homothallic fungus, meaning it is able to self-fertilize and form fruiting bodies in the absence of a mating partner. It has septate hyphae with a woolly colony texture and white mycelia. The green colour of wild-type colonies is due to pigmentation of the spores, while mutations in the pigmentation pathway can produce other spore colours.

Fungi are a diverse group of organisms that employ a huge variety of reproductive strategies, ranging from fully asexual to almost exclusively sexual species. Most species can reproduce both sexually and asexually, alternating between haploid and diploid forms. This contrasts with most multicellular eukaryotes such as mammals, where the adults are usually diploid and produce haploid gametes which combine to form the next generation. In fungi, both haploid and diploid forms can reproduce – haploid individuals can undergo asexual reproduction while diploid forms can produce gametes that combine to give rise to the next generation.
Sporogenesis is the production of spores in biology. The term is also used to refer to the process of reproduction via spores. Reproductive spores were found to be formed in eukaryotic organisms, such as plants, algae and fungi, during their normal reproductive life cycle. Dormant spores are formed, for example by certain fungi and algae, primarily in response to unfavorable growing conditions. Most eukaryotic spores are haploid and form through cell division, though some types are diploid sor dikaryons and form through cell fusion.we can also say this type of reproduction as single pollination
Pathogenic fungi are fungi that cause disease in humans or other organisms. Although fungi are eukaryotic, many pathogenic fungi are microorganisms. Approximately 300 fungi are known to be pathogenic to humans; their study is called "medical mycology". Fungal infections are estimated to kill more people than either tuberculosis or malaria—about two million people per year.
Homothallic refers to the possession, within a single organism, of the resources to reproduce sexually; i.e., having male and female reproductive structures on the same thallus. The opposite sexual functions are performed by different cells of a single mycelium.

Sexual reproduction is a type of reproduction that involves a complex life cycle in which a gamete with a single set of chromosomes combines with another gamete to produce a zygote that develops into an organism composed of cells with two sets of chromosomes (diploid). This is typical in animals, though the number of chromosome sets and how that number changes in sexual reproduction varies, especially among plants, fungi, and other eukaryotes.

Brachymeiosis was a hypothesized irregularity in the sexual reproduction of ascomycete fungi, a variant of meiosis following an "extra" karyogamy step. The hypothesized process would have transformed four diploid nuclei into eight haploid ones. The current scientific consensus is that brachymeiosis does not occur in any fungi.
References
- ↑ Alexopolous (1996), et al., pp. 196–97.
- 1 2 3 4 5 Deacon J. (2005). Fungal Biology. Cambridge, MA: Blackwell Publishers. pp. 167–68. ISBN 1-4051-3066-0.
- ↑ Sidhu GS. (1983). "Sexual and parasexual variability in soil fungi with special reference to Fusarium moniliforme". Phytopathology. 73 (6): 952–55. doi:10.1094/phyto-73-952.
- ↑ Durand N, Reymond P, Fevre M (1992). "Transmission and modification of transformation markers during an induced parasexual cycle in Penicillium roqueforti". Current Genetics. 21 (4–5): 377–83. doi:10.1007/bf00351698. S2CID 30871714.
- ↑ Alexopolous (1996), et al., p. 12.
- ↑ Pulhalla JE, Mayfield JE (1974). "The mechanism of heterokaryotic growth in Verticillium dahliae". Genetics. 76 (3): 411–422. PMC 1213075 . PMID 17248647.
- ↑ O'Garro LW, Clarkson JM (1992). "Variation for pathogenicity on tomato among parasexual recombinants of Verticillium dahliae". Plant Pathology. 41 (2): 141–47. doi:10.1111/j.1365-3059.1992.tb02331.x.
- ↑ Hastie AC. (1964). "The parasexual cycle in Verticillium albo-atrum". Genetics Research. 5 (2): 305–15. doi: 10.1017/s0016672300001245 .
- ↑ Hocart MJ, Lucas JA, and Peberdy JF. "Parasexual recombination between W and R pathotypes of Pseudocercosporella herpotrichoides through protoplast fusion." Mycological Research. 1993 August;97(8):977-983.
- ↑ Garber ED, Ruddat M (1992). "The parasexual cycle in Ustilago scabiosae (Ustilaginales)". International Journal of Plant Sciences. 153: 98–101. doi:10.1086/297010.
- ↑ Zeigler RS, Scott RP, Leung H, Bordeos AA, Kumar J, Nelson RJ (1997). "Evidence of parasexual exchange of DNA in the rice blast fungus challenges its exclusive clonality". Phytopathology. 87 (3): 284–94. doi: 10.1094/phyto.1997.87.3.284 . PMID 18945171.
- ↑ Higgins VJ, Miao V, Hollands J (1987). "The use of benomyl and cycloheximide resistance markers in studies of race development by the leaf mold pathogen Cladosporium fulvum". Canadian Journal of Plant Pathology. 9: 14–19. doi:10.1080/07060668709501905.
- ↑ Arnaru J, Oliver RP (1993). "Inheritance and alteration of transformed DNA during an induced parasexual cycle in the imperfect fungus Cladosporium fulvum". Current Genetics. 23 (5–6): 508–11. doi:10.1007/bf00312643. PMID 8319310. S2CID 25780981.
- ↑ Bennett RJ and Johnson AD. "Completion of a parasexual cycle in Candida albicans by induced chromosome loss in tetraploid strains." EMBO J. 2003 May 15;22(10):2505-15.
- ↑ Seervai RNH, Knox SKJ, Hirakawa MK, Porman AM, and Bennett RJ. "Parasexuality and Ploidy Change in Candida tropicalis." Eukaryotic Cell. 2013 Dec; 12(12): 1629–1640.
- 1 2 Butler G, Rasmussen MD, Lin MF, Santos MA, Sakthikumar S, Munro CA, et al. (June 2009). “Evolution of pathogenicity and sexual reproduction in eight Candida genomes”. Nature. 459 (7247): 657–662. Bibcode:2009Natur.459..657B. doi:10.1038/nature08064. PMC 2834264. PMID 19465905
Cited text
- Alexopoulos CJ, Mims CW, Blackwell M (1996). Introductory Mycology. John Wiley and Sons. pp. 196–97. ISBN 0-471-52229-5.
| Biological terms | |
|---|---|
| Sexual reproduction | |
| Sexuality | |